 О проектеПредмет клеточнойСтроение и химияЦитоплазмаСистемыЦитоскелетМеханизмы
О проектеПредмет клеточнойСтроение и химияЦитоплазмаСистемыЦитоскелетМеханизмыЦЕНТРАЛЬНАЯ ДОГМА МОЛЕКУЛЯРНОЙ БИОЛОГИИ
ХРОМОНЕМНЫЙ (ЧЕТВЕРТЫЙ) УРОВЕНЬ УПАКОВКИ ХРОМАТИНА
ЯДРЫШКО - ИСТОЧНИК РИБОСОМ
НЕРИБОСОМНЫЕ ПРОДУКТЫ КЛЕТОЧНОГО ЯДРА
ЯДЕРНАЯ ОБОЛОЧКА

Внутри интерфазных ядер как при витальных наблюдениях, так и на фиксированных и окрашенных препаратах видны мелкие, обычно шаровидные тельца - ядрышки. Впервые ядрышки были обнаружены Фонтана в 1774 г. В живых клетках они выделяются на фоне диффузной организации хроматина из-за своей светопреломляемости. Последнее свойство связано с тем, что ядрышки являются наиболее плотными структурами в клетке. Они обнаруживаются практически во всех ядрах эукариотических клеток за редким исключением. Это говорит об обязательном присутствии этого компонента в клеточном ядре.
В клеточном цикле ядрышко присутствует в течение всей интерфазы, в профазе по мере компактизации хромосом во время митоза оно постепенно исчезает и отсутствует в мета- и анафазе, вновь появляется в середине телофазы, чтобы сохраняться вплоть до следующего митоза, или до гибели клетки.
Долгое время функциональное значение ядрышка было непонятно. Вплоть до 1950-х годов исследователи считали, что вещество ядрышка представляет собой своего рода запас, который используется и исчезает в момент деления ядра.
Еще в 1930-х годах рядом исследователей (МакКлинток, Хейтц, С.Г. Навашин) было показано, что возникновение ядрышек связано топографически с определенными зонами на особых, ядрышкообразующих хромосомах. Эти зоны были названы ядрышковыми организаторами, а сами ядрышки представлялись как структурное выражение хромосомной активности. Позднее, в 1940-х годах, когда было найдено, что ядрышки содержат РНК, стала понятна их «базофилия», сродство к основным (щелочным) красителям вследствие кислой природы РНК. По данным цитохимических и биохимических исследований, основным компонентом ядрышка является белок: на его долю приходится до 70—80% от сухой массы. Такое большое содержание белка и определяет высокую плотность ядрышек. Кроме белка в составе ядрышка обнаружены нуклеиновые кислоты: РНК (5-14%) и ДНК (2-12%).
Уже в 1950-х годах при изучении ультраструктуры ядрышек в их составе были выявлены гранулы, сходные по своим свойствам с цитоплазматическими гранулами рибонуклеопротеидной природы — с рибосомами. Следующим этапом в изучении ядрышка было открытие принципиального факта — «ядрышковый организатор» является вместилищем генов рибосомных РНК.
Строение рибосом
Рибосома представляет собой элементарную клеточную машину синтеза любых белков клетки. Все они построены в клетке одинаково, имеют одинаковую молекулярную композицию, выполняют одинаковую функцию — синтез белка, поэтому их также можно считать клеточными органоидами. В отличие от других органоидов цитоплазмы (пластид, митохондрий, клеточного центра, мембранной вакуолярной системы и др.) они представлены в клетке огромным числом: за клеточный цикл их образуется 1·107 штук. Поэтому основная масса клеточной РНК представляет собой именно рибосомную РНК. РНК рибосом относительно стабильна, рибосомы могут существовать в клетках культуры ткани в течение нескольких клеточных циклов. В печеночных клетках время полужизни рибосом составляет 50-120 ч.
Рибосомы — это сложные рибонуклеопротеидные частицы, в состав которых входит множество молекул индивидуальных (неповторенных) белков и несколько молекул РНК. Рибосомы прокариот и эукариот по своим размерам и молекулярным характеристикам отличаются, хотя и обладают общими принципами организации и функционирования. К настоящему времени методом рентгеноструктурного анализа высокого разрешения полностью расшифрована структура рибосом.
Полная, работающая рибосома состоит из двух неравных субъединиц, которые легко обратимо диссоциируют на большую субъединицу и малую. Размер полной прокариотической рибосомы составляет 20 х 17 х 17 нм, эукариотической — 25 х 20 х 20. Полная прокариотическая рибосома имеет коэффициент седиментации 70S и диссоциирует на две субъединицы: 50S и 30S. Полная эукариотическая рибосома с коэффициентом седиментации 80S диссоциирует на субъединицы 60S и 40S. Форма и детальные очертания рибосом из разнообразных организмов и клеток, включая как прокариотические, так и эукариотические, поразительно похожи, хотя и отличаются рядом деталей. Малая рибосомная субъединица имеет палочковидную форму с несколькими небольшими выступами (рис. 81), ее длина составляет около 23 нм, а ширина — 12 нм. Большая субъединица похожа на полусферу с тремя торчащими выступами. При ассоциации в полную 70S рибосому малая субчастица ложится одним концом на один из выступов 50S частицы, а другим — в ее желобок. В состав малых субъединиц входит по одной молекуле РНК, а в состав большой — несколько: у прокариот — две, а у эукариот — три молекулы. Характеристики молекулярной композиции рибосом даны в табл. 9.
|
Рис. 81. Рибосомы бактерий в разных проекциях а — малая субъединица; 6 — большая субъединица; в — полная 70S рибосома (вид сверху и сбоку) |
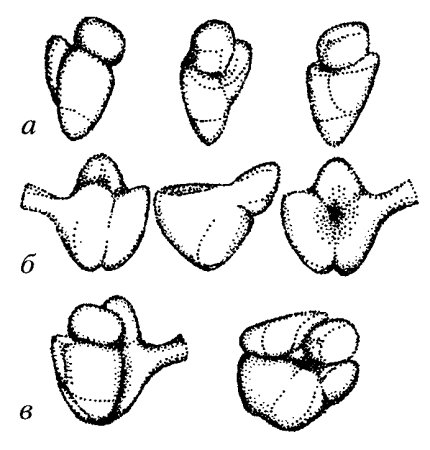
Таблица 9 |
Молекулярная характеристика рибосом |
|
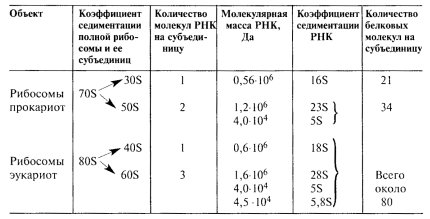
Таким образом, в состав эукариотической рибосомы входят четыре молекулы РНК разной длины: 28S РНК содержит 5000 нуклеотидов, 18S РНК - 2000, 5,8S РНК - 160, 5S РНК - 120. Рибосомные РНК обладают сложной вторичной и третичной структурой, образуя сложные петли и шпильки на комплементарных участках, что приводит к самоупаковке, самоорганизации этих молекул в сложное по форме тело. Например, сама по себе молекула 18S РНК в физиологических ионных условиях образует палочковидную частицу, определяющую форму малой субъединицы рибосомы.
Под действием низких ионных сил, особенно при удалении ионов магния, плотные рибосомные субъединицы могут разворачиваться в рыхлые рибонуклеопротеидные тяжи, где удается наблюдать кластеры отдельных белков, но правильных структур типа нуклеосом нет, так как нет групп из сходных белков: в рибосоме все 80 белков разные.
Для того чтобы образовались рибосомы, необходимо наличие четырех типов рибосомных РНК в эквимолярных отношениях и наличие всех рибосомных белков. Сборка рибосом может происходить спонтанно in vitro, если последовательно добавлять к РНК белки в определенной последовательности.
Следовательно, для биосинтеза рибосом необходим синтез множества специальных рибосомных белков и четырех типов рибосомной РНК. Где эта РНК синтезируется, на каком количестве генов, где эти гены локализованы, как они организованы в составе ДНК хромосом — все эти вопросы в последние десятилетия были успешно разрешены при изучении строения и функции ядрышек.
Чем определяется число ядрышек в клетке
Как уже говорилось, все клетки имеют обязательные внутриядерные структуры — ядрышки. Это правило имеет исключения, которые только подчеркивают важность и необходимость участия ядрышка и жизненных отправлениях клетки. К таким исключениям относятся клетки дробящихся яиц, где ядрышки отсутствуют на ранних этапах эмбриогенеза, или клетки, закончившие развитие и необратимо специализировавшиеся, например некоторые клетки крови.
В остальных случаях в клетках наблюдается 1—5 ядрышек, причем их количество не строго постоянно даже у одного и того же типа клеток. Более того, в некоторых половых клетках (растущие ооциты) число ядрышек может достигать нескольких сотен, т.е. на два порядка выше, чем в соседних соматических клетках. Увеличение числа ядрышек называется амплификацией ядрышек.
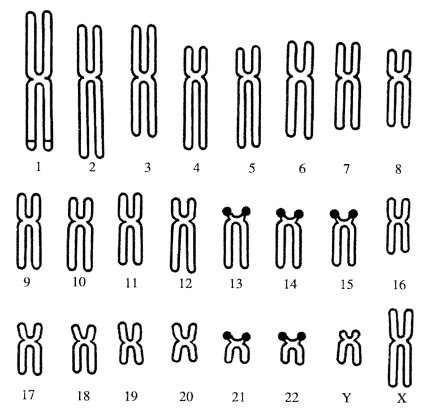
Еще в 1930-х годах было сделано предположение, что число ядрышек зависит от числа «ядрышковых организаторов» — особых участков, на которых в телофазе происходит новообразование ядрышек интерфазного ядра. Часто ядрышковые организаторы локализованы во вторичных перетяжках хромосом (образуют вторичные перетяжки хромосом). Так, у человека ядрышковые организаторы расположены в коротких плечах 13, 14, 15, 21 и 22-й хромосом (10 на диплоидный набор) (рис. 82). У млекопитающих обычно имеется несколько ядрышкообразующих хромосом на диплоидный набор: у кошки — 2; у свиньи — 2; у мыши — 4; у коровы — 8. У хладнокровных позвоночных и у птиц обычно имеется только по одной паре ядрышкообразующих хромосом.
|
Рис. 82. Хромосомы (13, 14, 15, 21, 22) с ядрышковыми организаторами в кариотипе человека |
Таким образом, максимальное число ядрышек в разных клетках определяется числом ядрышковых организаторов и увеличивается согласно плоидности ядра: в крупных полиплоидных ядрах всегда количество ядрышек больше.
Это правило подтверждается прямыми наблюдениями над мутантными особями с разным числом ядрышковых организаторов. Так, у шпорцевой лягушки в норме в диплоидной клетке есть две ядрышкообразующие хромосомы и соответственно 1-2 ядрышка. У гетерозиготной особи с одной ядрышкообразующей хромосомой имеется одно ядрышко, у гомозиготных мутантных личинок, у которых нет ядрышковых организаторов, ядрышки не возникают и не происходит синтез рРНК. Сходные наблюдения были получены на дрозофилах с разным числом ядрышкообразующих хромосом — от 0 до 4.
Локализация ядрышковых организаторов определяется довольно точно на митотических хромосомах с помощью окраски солями серебра, которые имеют специфическое сродство к некоторым ядрышковым белкам. Более точным является определение ядрышковых организаторов с помощью метода молекулярной гибридизации in situ. Так, меченная тритием рРНК при контакте с денатурированной ДНК на препарате митотических хромосом образует ДНК-рРНК-гибрид только в тех местах, где есть последовательности ДНК, комплементарные рРНК.
Чаще всего в клетках количество ядрышек меньше, чем число ядрышковых организаторов. Это связано с тем, что при новообразовании ядрышек они могут сливаться друг с другом в одну общую структуру, т.е. в пространстве интерфазного ядра отдельные ядрышковые организаторы разных хромосом могут объединяться. Так, в тканях человека могут встречаться клетки с одним ядрышком. Это значит, что десять ядрышкообразующих участков - локусов, диплоидного набора хромосом входят в состав одного ядрышка. Слияние ядрышек друг с другом хорошо показано на живых клетках культуры ткани при цейтраферной киносъемке.
Множественность рибосомных генов
При изучении числа ядрышек при различных хромосомных абберациях было найдено, что при разрыве хромосомы на месте вторичной перетяжки ядрышки могут возникать на каждом из фрагментов хромосом. Так, при обмене участками между двумя хромосомами в микроспороцитах кукурузы, в том случае, когда разрыв одной из хромосом происходил через ядрышковый организатор, возникали две хромосомы, каждая из которых несла часть исходного ядрышкового организатора. В этом случае обе хромосомы обладали способностью образовывать ядрышки, хотя и в неодинаковой степени. Из этих наблюдений был сделан очень важный вывод (который полностью подтвердился н 1960-х годах на молекулярно-биологическом уровне) о том, что ядрышковый организатор представляет собой не точечный локус хромосомы, а является множественным по своей структуре, содержит несколько одинаковых генных участков, каждый из которых отвечает за образование ядрышка.
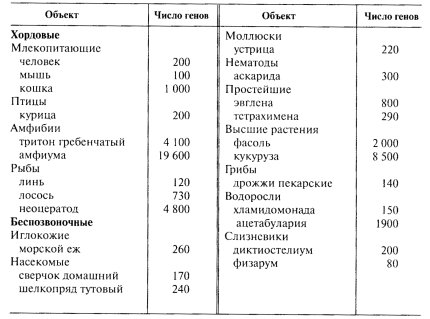
Методом молекулярной гибридизации было показано, что в составе геномов эукариот рибосомные гены представлены сотнями и тысячами единиц; они принадлежат к фракции умеренно повторяющихся последовательностей ДНК. Даже у бактерий в геноме может быть несколько (6—7) рассеянных по геному идентичных последовательностей, ответственных за синтез рРНК. Общее количество этой фракции ДНК (рДНК) у Е. coli составляет около 1% от всей ДНК. У эукариотических организмов этот процент может составлять 0,18 для X. laevis, 0,4 - для человека, 1,3 - для дрозофилы и 5,5% - для пекарских дрожжей. Число рибосомных генов у эукариот намного больше, чем у прокариотических клеток. В табл. 10 приведены некоторые примеры числа генов рРНК у различных представителей эукариот.
Таблица 10 |
Количество рибосомных генов на гаплоидный набор хромосом |
|
С помощью метода молекулярной гибридизации проанализировано не только число рибосомных генов, но и их локализация. Из этих экспериментов следовало, что именно зоны ядрышковых организаторов во вторичных перетяжках хромосом Xenopus содержат рибосомные гены и что в каждом из этих организаторов имеется примерно по 300 генов, т.е. ядрышковые организаторы представляют собой полицистронные участки, содержащие множество одинаковых генов (полиизогенные участки). Следовательно, рибосомные гены собраны вместе в группы, или кластеры.
Наблюдать непосредственно порядок расположения рибосомных генов на ДНК выделенных ядрышек с помощью электронного микроскопа удалось на дополнительных ядрышках ооцитов амфибий.
Амплифицированные ядрышки
Обычно число генов рибосомных РНК постоянно на геном, оно не меняется в зависимости от уровня транскрипции этих генов. Например, у клеток с высоким уровнем метаболизма число генов рРНК точно такое же, как у клеток, полностью прекративших синтез рибосом. При репликации ДНК в S -периоде происходит и удвоение числа генов рРНК, поэтому их количество коррелирует с плоидностью клетки.
Однако существуют случаи, когда гены рРНК подвергаются избыточной репликации. При этом дополнительная репликация генов рРНК происходит в целях обеспечения продукции большого количества рибосом. В результате такого сверхсинтеза генов рРНК их копии могут становиться свободными, экстрахромосомными. Эти внехромосомные копии генов рРНК могут функционировать независимо, в результате чего возникает масса свободных дополнительных ядрышек, но уже не связанных структурно с ядрышкообразующими хромосомами. Это явление получило название амплификации генов рРНК. Особенно подробно это явление изучено на растущих ооцитах амфибий, хотя оно встречается как у животных, так и у растений.
Так, у X. laevis (наиболее подробно изученный и популярный объект) амплификация рДНК происходит в профазе I деления созревания, когда синтез хромосомной ДНК давно закончен. В этом случае количество амплифицированной рДНК (или генов рРНК) становится в 3000 раз больше того, что приходится на гаплоидное количество рДНК, и соответствует 1,5·106 генов рРНК. Эти сверхчисленные внехромосомные копии и образуют сотни дополнительных ядрышек в растущих ооцитах. В среднем же на одно дополнительное ядрышко приходится несколько сотен или тысяч генов рРНК.
Амплифицированные ядрышки встречаются также в ооцитах насекомых. Так, у окаймленного плавунца в ооцитах обнаружено 3·106 экстрахромосомных копий генов рРНК.
Биологический смысл появления сверхчисленных экстрахромосомных ядрышек при росте ооцитов вполне понятен: для синтеза огромного количества запасных продуктов, которые будут использованы на ранних стадиях эмбриогенеза, необходимо соответственно огромное количество рибосом, которые могут быть синтезированы в клетке на дополнительных матрицах этих многочисленных амплифицированных ядрышек. После периода созревания ооцита при его двух последовательных делениях эти дополнительные ядрышки в состав митотических хромосом не входят, они отделяются от новых ядер и деградируют. Следовательно, амплификация рДНК в ооците представляет собой временное явление, не сказывающееся на постоянстве генома.
У низших эукариотических организмов наблюдаются также экстрахромосомные ядрышки. Например, у Tetrachymena pyriformis в составе гаплоидного генома микронуклеуса имеется только единственный ген рРНК. В макронуклеусе этого организма содержится около 200 гаплоидных эквивалентов в виде экстрахромосомных копий. У дрожжевых клеток также обнаружены экстрахромосомные копии генов рРНК в виде циклических молекул ДНК длиной около 3 мкм, содержащих один ген рРНК.
Строение и функционирование генов рРНК
Итак, в ядрышковых организаторах определенных хромосом локализованы места множественных сгруппированных вместе генов рибосомной РНК. Но как уже говорилось, существуют четыре типа молекул рибосомной РНК, каждый из которых в полной эукариотической рибосоме представлен один раз. Значит ли это, что для каждой из этих РНК (28S рРНК, 18S рРНК, 5,8S рРНК, 5S рРНК) должен существовать отдельный ген, долгое время было неясным. Непонятным было также, как осуществляется в клетках одновременное сбалансированное образование этих разных рРНК. Этот вопрос был решен при исследовании динамики синтеза рибосомных РНК. Обнаружено, что при использовании импульсной короткой метки среди клеточных РНК выявляется быстро синтезирующая РНК с высокой скоростью седиментации — тяжелая 45S РНК. Если после появления этой 45S РНК продолжать наблюдать за распределением метки во фракциях РНК, но уже в отсутствие меченых предшественников, то можно видеть, что по мере убывания метки в зоне 45S РНК она начинает появляться и стабильно накапливаться в зонах 28S, 18S и 5,8S рибосомных РНК. Эти данные говорили о том, что при синтезе рибосомных РНК сначала образуется гигантская молекула-предшественник (45S РНК), которая затем дает начало основным молекулам рибосомной РНК. Найдено, что молекула 45S РНК содержит около 13·103 оснований, имеет массу около 4,6·106 и может достигать 2—5 мкм в длину. Явление распада молекулы 45S рРНК на фрагменты, соответствующие размерам 28S, 18S и 5,8S РНК, получило название «процессинг», или созревание. Во время процессинга происходит разрыв предшественника на три фрагмента и, кроме того, наблюдается значительная деградация РНК (около 50%, т. е. 6 000 нуклеотидов). Кроме этих данных было вычислено, что молекула 5S РНК синтезируется независимо от 45S РНК и локализация гена 5S рРНК не связана с ядрышковым организатором.
Почти одновременно с получением этих биохимических данных О. Миллеру (1969) удалось с помощью электронного микроскопа увидеть работающие рибосомные гены. Для этого под световым микроскопом вручную выделили ядра из средних ооцитов тритона, микроиглами разорвали ядерную оболочку и в микропипетку втянули многочисленные амплифицированные ядрышки. Такая капля, содержащая ядрышки и кариоплазму, была перенесена в раствор низкой ионной силы со щелочным значением среды. Этот раствор наслаивали на раствор сахарозы с формалином, находящийся в микроячейке центрифужной пробирки, на дно микроячейки помещали сеточку с формваром для электронной микроскопии. Действие низкой ионной силы в щелочной среде приводило к набуханию и диспергированию выделенных ядрышек, они разрыхлялись настолько, что становились плохо различимыми в световом микроскопе. При центрифугировании такие набухшие ядрышки проходили через слой сахарозы, еще больше расправлялись и фиксировались в формалине. Наконец, они достигали дна микроячейки и распластывались на формваровой подложке. После этого сеточки вынимали, обезвоживали, оттеняли металлом и просматривали в электронном микроскопе (рис. 83).
|
Рис. 83. Рибосомные транскрипты в выделенных и деконденсированных дополнительных ядрышках из ооцитов тритона 1 — транскрибируемый участок (р-ген); 2 — спейсеры |
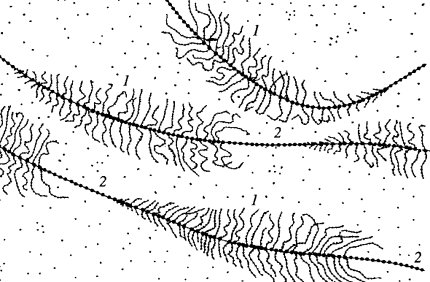
На таком препарате были видны сложно изогнутые и перепутанные длинные осевые молекулы ДНК, на которых через равные промежутки располагались фибриллярные зоны, имеющие вид «елочек». Длина фрагмента ДНК, занятого такой «елочкой», была постоянной и равнялась 5 мкм. На этом отрезке располагалось около 100 плотных гранул величиной около 20 нм, от каждой из которых отходила в сторону тонкая изогнутая нить. Величина такой нити была минимальной на одном конце такого отрезка и максимальной на другом. Эти извитые латеральные нити и образовывали структуру типа «елочки». Доказано, что крупные гранулы на нити ДНК представляют собой молекулы РНК-полимеразы I, ответственной за синтез рРНК, а боковые изогнутые нити — транскрипты, состоящие из синтезируемых молекул РНК. Самые длинные транскрипты находились на одном конце «елочки» и соответствовали 45S предшественнику рРНК. Следовательно, синтез рРНК начинался на конце отрезка с короткими боковыми нитями и заканчивался на участке с длинными нитями РНК. Такой участок ДНК, на котором были видны молекулы рРНК в процессе их удлинения, получил название транскрипционной единицы. Между транскрипционными единицами располагались участки ДНК, лишенные гранул РНК-полимеразы I и транскриптов. Это — зоны так называемых спейсеров, которые не транскрибируются, и, более того, на таких препаратах они имеют нуклеосомное строение, тогда как транскрипционные единицы свободны от нуклеосом. Величина таких спейсерных участков может варьировать не только в данной клетке, но быть различной у разных видов. Длина боковых фибрилл была в 5—10 раз короче, чем 45S РНК, из-за того, что эта новосинтезированная РНК связана с белками, образуя рибонуклеопротеидный тяж — предшественник рибосом.
Исходя из этих работ стало очевидным то, что рибосомный ген состоит из двух участков: нетранскрибируемой последовательности ДНК (nts) — спейсера — и транскрипционной единицы. В состав транскрипционной единицы входят участки, соответствующие 28S, 18S и 5,8S рРНК, разделенные вставками, которые деградируют при процессинге 45S РНК.
Расшифровка структуры рибосомных генов различных эукариотических объектов показала удивительно универсальный тип их строения:
3' nts—промотор—tse—18S pPHK—tsi1—5,8S pPHK—tsi2—28S рРНК 5',
где nts — нетранскрибируемые последовательности ДНК спейсерного участка, ts - транскрибируемые последовательности ДНК (внешняя и две внутренние) и участки, соответствующие зрелым рибосомным РНК. В состав транскрипционной единицы входит весь ген за исключением спейсерного участка. Такая структура рибосомного гена практически одинакова для всех эукариотических организмов (рис. 84). Вариабельными являются как нетранскрибируемые (спейсерные) участки, так и транскрибируемые вставки (ts), которые не входят в состав зрелых рРНК.
|
Рис. 84. Строение р-генов разных организмов (А. Хаджиолов) tSe и tSi — соответственно внешняя и внутренние транскрибируемые последовательности |
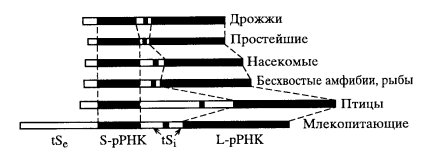
Итак, три основные молекулы рРНК синтезируются на одной транскрипционной единице. Что же касается молекулы 5S рРНК, то она к этому гену никакого отношения не имеет: 5S рРНК синтезируется на отдельных генах, локализованных не в зонах ядрышковых организаторов, а на совсем иных хромосомах при участии РНК-полимеразы III. Так, у человека основная масса генов 5S рРНК находится на 1 хромосоме, более мелкие кластеры — на 9 и 16 хромосомах. У ксенопуса гены 5S рРНК расположены в теломерных участках большинства хромосом. Гены 5S рРНК, тоже множественные, также собраны в кластеры, но их число выше, чем у остальных генов рРНК. Например, у человека их насчитывается до 2000, у ксенопуса - 24 000, у гребенчатого тритона - 32 000. Есть объекты и с меньшим их числом: дрозофила - 320, крыса - 830. У нейроспоры и дрожжей число генов 5S рРНК одинаковое с числом других рибосомных генов, так как участок 5S рРНК включен и транскрибируется в спейсерной зоне.
Транскрипция рРНК идет с помощью двух ферментов: РНК-полимеразы I, которая участвует в синтезе 45S предшественника рРНК и РНК-полимеразы III, ответственной за синтез 5S рРНК. Матрицей для синтеза рРНК по определению должна быть ядрышковая ДНК.
В изолированном р-хроматине обнаружены гистоны, негистоновые белки и белки рибосом. Так, в р-хроматине выявлены основные сердцевинные (кóровые) гистоны, но их количество составляет только 40% по сравнению с таковым в тотальном хроматине.
Первичные транскрипты (морфологически представлены в виде латеральных филаментов на «елочках», образующихся на активных транскрипционных единицах) прогрессивно увеличиваются в длину по мере прохождения РНК-полимеразы I вдоль всего транскрипционного участка гена, начиная с точки начала репликации до терминального участка. Скорость роста цепи пре-рРНК составляет около 20-30 нуклеотидов в секунду, т.е. весь синтез 45S рРНК. занимает около 5-10 мин.
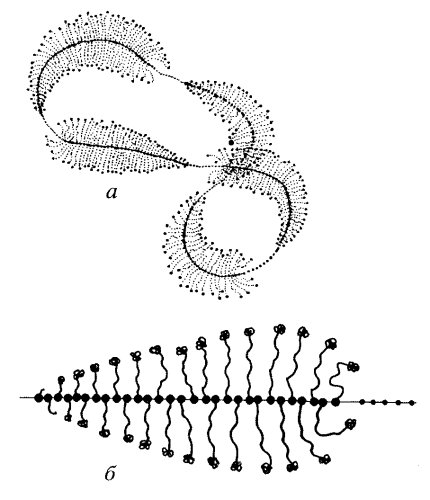
На каждой транскрипционной единице располагается множество (50-100) молекул РНК-полимеразы I, тем самым на каждом гене одновременно происходит синтез множества молекул пре-рРНК, которые находятся на разных стадиях роста полинуклеотидной цепи (рис. 85). Максимальную величину пре-рРНК имеет вблизи терминального участка, где ее молекулярная масса достигает 4,5·106 Да (для млекопитающих), а длина должна соответствовать 5,2 мкм. На самом же деле длина латерального транскрипта в 5—10 раз короче этой величины. Это обусловлено тем, что по мере роста транскрипта он связывается сразу же с белками, образуя в конечном участке транскрипции рибонуклеопротеид с коэффициентом седиментации 80S. Такие 80S рРНП составляют до 20% от всех РНП ядрышка. Большая часть белков, которые связываются с 45S РНК, являются белками, входящими в состав малой и большой субъединиц зрелых рибосом. Таким образом, уже на уровне незрелой гигантской молекулы пре-рРНК происходит специфическое связывание с рибосомными белками: около 50% белков большой субъединицы и около 30% малой субъединицы связываются с пре-рРНК во время ее синтеза или вскоре после него. Такая связь 45S РНК с белками приводит к тому, что латеральные транскрипты имеют толщину около 10 нм (после оттеснения металлами), на их свободном конце часто наблюдается крупная гранула (30 нм), что может указывать на высокую степень компактизации РНК и белка на 5'-конце цепи РНК.
|
Рис. 85. Транскрипция р-генов на циклической ДНК дополнительного ядрышка ооцита жука-плавунца (а) и строение одной транскрипционной единицы (р-гена) (б) |
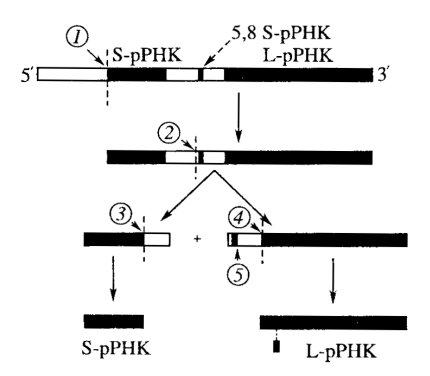
Распад 45S РНК на более короткие отрезки, явление созревания рРНК, или процессинг, происходят после завершения транскрипции. Ферментативный механизм этого явления до конца еще не ясен, в нем принимают участие эндо- и экзонуклеазы. При этом происходят последовательное расщепление пре-рРНК на фрагменты и частичная деградация участков РНК на этих фрагментах. В результате процессинга пре-рРНК примерно 50% нуклеотидов первично синтезированной молекулы отщепляется (мол. масса 45S РНК составляет 4,6·106 Да, а суммарная мол. масса зрелых рРНК - около 2,2·106 Да) (рис. 86).
|
Рис. 86. Процессинг 45S РНК 1—5 — места последовательного расщепления предшественника рибосомных РНК |
Таким образом, в ядрышке локализуются следующие основные предшественники рибосом: 1) транскрипты рРНК в процессе их роста; 2) 80S РНП, содержащие 45S РНК, могут составлять до 10-20% всех РНП ядрышка; 3) 55S РНП - предшественники большой субъединицы, могут составлять до 70-80% всех РНП ядрышка; время созревания большой рибосомной субъединицы занимает около одного часа; 4) незрелые малые (40S РНП) субъединицы рибосом, быстро (за 15—30 мин) покидающие ядрышко.
В интенсивно функционирующих ядрышках происходит синтез огромного числа рибосом: 1500-3000 штук в минуту. Поэтому в ядрышке насчитывается около 5·104 предшественников рибосом.
Структура ядрышка
Сведения о тонком строении ядрышка были получены главным образом методом электронной микроскопии. Световая микроскопия давала ограниченный набор данных о структуре ядрышка из-за малого их размера (1-5 мкм) и недостаточной разрешающей способности этого метода. Из прижизненных наблюдений видно, что ядрышки обладают высокой плотностью и высоким светопреломлением. В их структуре даже прижизненно видна некоторая неоднородность: нитчатые (нуклеолонемы), гранулярные компоненты (нуклеолини), а также светлые зоны - «вакуоли». Гистохимически в ядрышках выявлялась РНК, но не ДНК. ДНК обнаруживалась лишь в периферической зоне ядрышек в виде так называемого околоядрышкового хроматина который мог прилежать к одной из сторон ядрышка, окружать его кольцом или вообще отсутствовать. Считалось, что околоядрышковый хроматин представляет собой гетерохроматиновые зоны. Кроме того было найдено, что ядрышки имеют некоторое сродство к солям серебра, т.е. обладают аргентофилией, могут восстанавливать серебро из различных растворов: нитрата серебра, «аммиачного серебра», протеинатов серебра. При этом происходит отложение темных осадков исключительно в ядрышках интерфазных клеток, а также в ядрышковых организаторах на митотических хромосомах при делении клетки.
Первые электронно-микроскопические работы показали, что ядрышки самых различных объектов, несмотря на их разнообразие, построены из одинаковых компонентов: гранулярного и фибриллярного (рис. 87). При этом гранулы в составе ядрышек имели размеры 15- 20 нм и были несоизмеримо меньше тех «гранул», что были видны в световом микроскопе. Кроме гранул в составе ядрышек обнаружили зоны скопления тонких (3-5 нм) фибрилл - диффузная часть ядрышек. Взаимное расположение гранулярных и фибриллярных зон в ядрышке может быть различным. Так, в некоторых случаях фибриллярный компонент занимает центральную часть ядрышка в виде однородного образования (печень аксолотля, многие ядрышки растительных меристем) или в виде нескольких (3-5) отдельных зон (рис. 88).
|
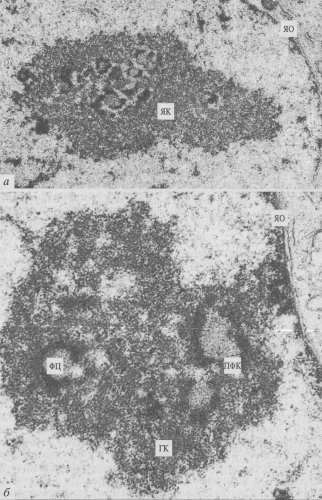
Рис. 87. Строение ядрышка клеток культуры СПЭВ при малом (а) и большом (б) увеличении (фото О.В. Зацепиной) ЯО — ядерная оболочка; ЯК — ядрышко; ГК — гранулярный компонент; ФЦ — фибриллярный центр; ПФК — плотный фибриллярный компонент |
|
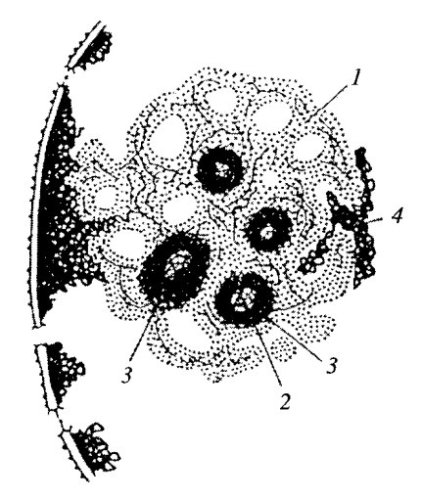
Рис. 88. Схема компонентов ядрышка 1 — гранулярный компонент (нуклеолонема); 2 — фибриллярные центры; 3 — плотный фибриллярный компонент; 4 — околоядрышковый хроматин |
Обычно гранулярный компонент (ГК) расположен на периферии ядрышка, но встречаются случаи, когда фибриллярный и гранулярный компоненты распределены в ядрышке равномерно. Часто в структуре ядрышек фибриллярно-гранулярные компоненты образуют нитчатые структуры - нуклеолонемы (ядрышковые нити), толщиной около 100—200 нм. Эти нуклеолонемы при достаточном контрастировании могут быть видны даже в световом микроскопе. Ядрышковые нити, или нуклеолонемы, также неоднородны по своему строению: в них кроме гранул толщиной 15 нм входит множество тонких фибрилл, которые могут образовывать в нуклеолонемах отдельные сгущения.
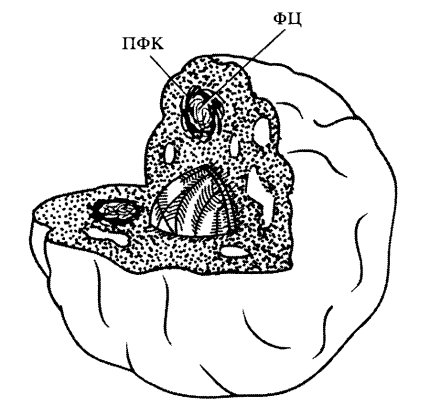
Неоднородной оказалась структура и диффузного, фибриллярного компонента. Найдено, что практически во всех типах ядрышек как животных, так и растительных объектов встречаются так называемые фибриллярные центры (ФЦ) - участки скопления фибрилл с низкой электронной плотностью, окруженные зоной фибрилл более высокой электронной плотности — плотный фибриллярный компонент (ПФК).
Кроме гранул и фибриллярных участков в структуре ядрышка обнаруживаются хроматиновые компоненты: такие как околоядрышковый хроматин, который может примыкать к ядрышку и даже окружать его. Часто 30-нанометровые фибриллы хроматина по периферии ядрышка заходят в лакуны между нуклеолонемными участками.
Наконец, в составе ядрышка выявляется белковый остов — матрикс. На ультратонких срезах необработанных ядрышек матрикс не выявляется в виде отдельного компонента, но если экстрагировать из ядрышек РНК, ДНК и белки, связанные с ними, то можно видеть, что ядрышко как таковое не распадается, не теряет своей общей формы. После таких обработок структура ядрышка представлена рыхлой фибриллярной сетью, заполняющей объем ядрышка.
Таким образом, в структуре ядрышек можно различить следующие пять компонентов: гранулярный, фибриллярные центры, плотный фибриллярный компонент, хроматин, белковый сетчатый матрикс.
Каким же образом распределены внутри ядрышек рДНК, рРНК и белки, где располагаются матрицы для синтеза рРНК, где первичные транскрипты, где предшественники рибосом, зрелые рибосомы — все эти вопросы были решены с применением самых разнообразных молекулярно-биологических и цитологических методов. Одним из этих методов является метод регрессивного окрашивания нуклеиновых кислот. Он основан на том, что ионы уранила, связанные с ДНК, более легко вымываются со срезов при обработке их хелатоном ЭДТА, чем ионы, связанные с РНК. Это позволяет различить в ядре плотные окрашенные структуры, содержащие РНК, и структуры, потерявшие окраску, те, что содержат ДНК. Так, в разнообразных ядрах участки хроматина, как конденсированного, так и диффузного, теряют окраску, а компоненты, содержащие РНК, - сохраняют. В ядре при этом контрастно выделяются разнообразные РНП, содержащиеся в основном объеме ядра и ядрышка. При этом в ядрышках интенсивно окрашены многочисленные гранулы, они окрашены так же, как рибосомы цитоплазмы. Окрашенным является плотный фибриллярный компонент, фибриллярные центры окрашены слабее, а внутриядрышковый и околоядрышковый хроматины выглядят светлыми. Следовательно, можно предположить, что как гранулярный компонент, который скорее всего представляет субъединицы рибосом, так и плотный фибриллярный компонент содержат РНК.
Так, при короткой пульсовой метке тритированным уридином (3Н-уридин) первые следы мечения обнаруживались сначала (через 1 — 15 мин) в плотном фибриллярном компоненте (ПФК), а затем (до 30 мин) меченым оказывался гранулярный компонент (ГК). Важно отметить, что в фибриллярных центрах (ФЦ) метка не обнаруживалась. Из этого наблюдения был сделан вывод, что 45S пре-рРНК синтезируется в области плотного фибриллярного компонента, а гранулярный компонент ядрышка соответствует прерибосомным частицам (55S-, 40S РНП).
Оставался открытым вопрос о природе фибриллярных центров, окруженных плотными РНК-содержащими фибриллами. Различными методами (специфическое окрашивание с помощью осмий-амина, ДНКазы, меченной золотом, связыванием меченого актиномицина, прямой молекулярной гибридизацией с меченой рДНК) обнаружено, что в составе фибриллярных центров находится ДНК, ответственная за синтез рРНК. Зоны фибриллярных центров отличаются от остального хроматина тем, что состоят из тонких хроматиновых фибрилл, значительно обедненных гистоном H1 (что показано с помощью меченных коллоидным золотом антител).
Эти исследования позволили связать друг с другом данные молекулярной организации транскрибируемых рибосомных генов с данными морфологии ядрышек и выяснить топологию в объеме ядрышка процесса синтеза рибосомной РНК и образования рибосом.
По модели, предложенной Жоссеном (1984), в фибриллярных центрах расположены неактивные рибосомные гены и, возможно, спейсерные участки. Транскрипция пре-рРНК происходит по периферии фибриллярных центров, где плотный фибриллярный компонент представляет собой 45S пре-рРНК, располагающиеся в виде «елочек» на деконденсированных участках рДНК (рис. 89). После завершения транскрипции 45S РНК теряет связь с транскрипционной единицей на ДНК в зоне плотного фибриллярного компонента, каким-то еще непонятным образом переходит в гранулярную зону, где и происходят процессинг рРНК, образование и созревание рибосомных субъединиц.
|
Рис. 89. Трехмерная реконструкция ядрышка и одного фибриллярного центра Транскрипты рРНК соответствуют плотному фибриллярному компоненту (ПФК). рДНК, «спейсеры» и неактивные цистроны расположены внутри фибриллярного центра (ФЦ) |
Фибриллярный центр и ядрышковый организатор
Строение и химические характеристики фибриллярных центров (ФЦ) оказались практически одинаковыми с таковыми ядрышковых организаторов митотических хромосом. И те и другие построены из тесно ассоциированных фибрилл толщиной 6—10 нм и обладают характерной особенностью — окрашиваются солями серебра, что зависит от наличия особых ядрышковых белков, а также содержат РНК- полимеразу I. Однако число ФЦ в интерфазных ядрышках не соответствует числу ядрышковых организаторов в митозе. Так, в клетках культуры СПЭВ число ФЦ может быть в 2—4 раза выше, чем число ядрышковых организаторов (табл. 11).
Таблица 11 |
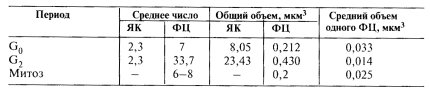
Количество ядрышек (ЯК), фибриллярных центров (ФЦ) в G0- и G2-периодах и во время митоза |
|
Более того, количество ФЦ возрастает по мере увеличения плоидности клетки (G2, 4n) и транскрипционной ее активности. При этом уменьшается величина каждого отдельного фибриллярного центра. Однако суммарные объемы ФЦ при пересчете на гаплоидный хромосомный набор остаются постоянными в интерфазе, но превышают это число вдвое по сравнению с метафазой. Другими словами, при активации синтеза рРНК наблюдается такое изменение числа ФЦ и их размеров, которое может говорить о какой-то фрагментации исходных ФЦ в относительно малоактивных ядрышках.
Противоположная картина наблюдается при затухании синтетических процессов в дифференцирующихся клетках эритроидного ряда мышей (табл. 12). При этом видно, что в размножающихся и активно синтезирующих гемоглобин проэритробластах количество фибриллярных центров зависит от плоидности клетки (88 в G1-фазе и 118 в G2-фазе клеточного цикла), размер индивидуальных ФЦ изменяется мало. После прекращения размножения этих клеток и падения их синтетической активности резко меняются параметры ядрышка. Их объем, начиная со стадии базофильного эритробласта, уменьшается в 4—5 раз, а на конечной стадии дифференцировки (нормобласт) — в сотню раз. При этом резко падает число ФЦ (10-40 раз) и почти в 10 раз возрастает величина отдельного фибриллярного центра.
Таблица 12 |
Количество фибриллярных центров и значения их размеров при эритропоэзе в печени зародышей мыши (приводятся средние значения) |
|
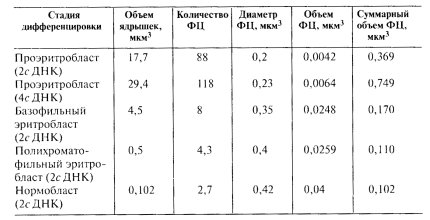
Исходя из этих наблюдений, можно так представить общую схему активации и инактивации ядрышка (рис. 90) на примере одного ядрышкового организатора. В неактивной форме ядрышковый организатор представлен в виде одного крупного фибриллярного центра, включающего в себя компактно уложенную часть цепи хромосомной ДНК, несущей тандемно расположенные рибосомные гены (транскрипционные единицы). В начале активации ядрышка происходит деконденсация р-генов на периферии такого фибриллярного центра. Эти р-гены начинают транскрибироваться, на них образуются РНП-транскрипты, которые при созревании дают начало появлению гранул — предшественников рибосом по периферии активированного ядрышка. По мере усиления транскрипции единый фибриллярный центр как бы распадается на ряд более мелких фибриллярных центров, связанных друг с другом полностью декомпактизованными участками рДНК. Чем выше транскрипционная активность ядрышка, тем больше число мелких, связанных друг с другом фибриллярных центров, окруженных плотным фибриллярным компонентом (ПФК), содержащим 45S рРНК. При полной активации ядрышка все мелкие фибриллярные центры деконденсируются; в этом случае зоны плотного фибриллярного компонента содержат всю рДНК, находящуюся в активном состоянии. Такая структура наблюдается у амплифицированных ядрышек растущих ооцитов. В случае инактивации ядрышка происходит постепенная конденсация рДНК, снова образуются фибриллярные центры, они объединяются друг с другом, величина их растет параллельно уменьшению доли ПФК. При полной инактивации, как в случае нормобластов, ядрышко представлено одним крупным (4-5 мкм) сферическим ФЦ без сопутствующего транскрипции ПФК: оно окружено зоной конденсированного хроматина. Такое инактивированное ядрышко сходно по своим структурным особенностям с ядрышковым организатором в составе митотических хромосом.
|
Рис. 90. Схема активации и инактивации ядрышкового организатора 1 — начало активации — «елочки» появляются по периферии одного крупного ФЦ (ЯОР-хромосомы); 2, 3 — дальнейшая активация приводит к образованию множественных ФЦ; 4 — полная активация; 5 — полная инактивация |
Структурные типы ядрышек
Приведенные выше описания дают основу для понимания разнообразия строения ядрышек в клетках с соответствующим уровнем синтеза рРНК. Однако кроме различной степени выраженности гранулярного и фибриллярных компонентов существуют и иные варианты структурной организации ядрышек. Обычно различают несколько структурных типов ядрышек: ретикулярный, или нуклеолонемный, компактный, кольцевидный, остаточный (покоящийся), сегрегированный (рис. 91).
|
Рис. 91. Структурные типы ядрышек 1 — ретикулярный; 2 — компактный; 3 — вакуолярный; 4 — кольцевидный; 5 — сегрегированный |
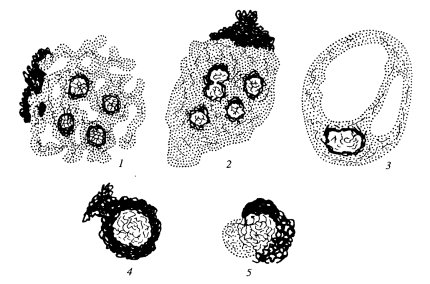
Ретикулярный тип ядрышка наиболее характерен для большинства клеток, для него свойственно нуклеолонемное строение, обилие гранул и фибриллярного плотного материала. Во многих случаях фибриллярные центры выявляются плохо, вероятно, из-за высокого уровня транскрипции. Этот тип ядрышек встречается в клетках животных и растений. Так, ретикулярный тип ядрышка гигантских политенных хромосом двукрылых насекомых и гигантских хромосом антиподиальных клеток ячменя очень сходен.
Компактный тип ядрышка отличается от предыдущего меньшей выраженностью нуклеолонемы, большей частотой встречаемости фибриллярных центров. Такие ядрышки характерны для активно размножающихся клеток (клетки растительных меристем, клетки культуры ткани и др.). Вероятно, оба эти типа могут переходить друг в друга, во всяком случае, они чаще всего встречаются в клетках с высоким уровнем синтеза РНК и белка.
Кольцевидные ядрышки встречаются в клетках животных. В световом микроскопе они имеют форму кольца с оптически светлой центральной зоной - это фибриллярный центр, окруженный РНП-фибриллами и гранулами. Размер этих ядрышек составляет около 1 мкм. Типичные кольцевидные ядрышки характерны для лимфоцитов и эндотелиоцитов, т.е. для клеток с относительно низким уровнем транскрипции.
Остаточные ядрышки характерны для клеток, полностью потерявших способность к синтезу рРНК (нормобласты, дифференцированные энтероциты, клетки шиповатого слоя кожного эпителия и др.). Часто они настолько малы и так окружены конденсированным хроматином, что с трудом обнаруживаются в световом микроскопе. В ряде случаев они могут снова активироваться и переходить в компактную или ретикулярную форму.
Сегрегированные ядрышки характерны для клеток, обработанных различными антибиотиками или химическими веществами, вызывающими прекращение синтеза рРНК (актиномицин Д, амфотерицин и др.), а также антибиотиками, влияющими на синтез ДНК и белков (митомицин, пуромицин, многие канцерогены и т.д.). Термин «сегрегация» используется в данном случае в связи с тем, что происходит как бы разделение, обособление разных компонентов ядрышек, сопровождающиеся прогрессивным уменьшением его объема. При этом обособляются друг от друга крупные фибриллярные центры и гранулярно-фибриллярный компонент.
Белки ядрышек
До 60% сухой массы выделенных ядрышек приходится на белки, число которых может составлять несколько сотен разных видов. Помимо белков ассоциированного с ядрышками хроматина в состав ядрышек входят белки рибосом и специфические ядрышковые белки, связанные с транскрипцией рибосомных генов, с процессингом 45S рРНК, такие как РНК-полимераза I, факторы транскрипции, топоизомеразы, метилазы, нуклеазы, протеинкиназы, фосфатазы. Часть ядрышковых белков имеет сродство к серебру (аргентофильные белки): РНК-полимераза I, фактор транскрипции UBF, нуклеолин (С-23), нуклеофозмин (ньюматрин, или В-23).
Аргентофилия характерна для белков, обогащенных сульфгидрильными, дисульфидными связями. Как уже указывалось, четкой аргентофилией обладают интерфазные ядрышки и зоны ядрышковых организаторов на митотических хромосомах.
Собственно ядрышковые белки расположены в специфических местах их активности. Так, РНК-полимераза I и фактор транскрипции рРНК UBF располагаются в фибриллярных центрах (ФЦ) и/или в плотном фибриллярном компоненте (ПФК).
Аргентофильным является также белок с молекулярной массой 195 кДа, представляющий собой большую субъединицу РНК-полимеразы I, участвующую в синтезе рРНК. Этот белок локализуется в зоне фибриллярных центров, по их периферии. На плоскостных препаратах ядрышек аргентофилией обладают участки над осевой частью «елочек», непосредственно над расположением гранул РНК-полимеразы I. Кроме того, с помощью иммуноморфологических методов РНК-полимераза I обнаруживается в зоне ядрышковых организаторов митотических хромосом. Это обстоятельство не противоречит данным о том, что во время митоза транскрипция полностью прекращается. Вероятно, что во время митоза гены, нагруженные неактивной РНК-полимеразой I, переносятся вместе с нею в области ядрышковых организаторов из одной клеточной генерации в другую.
Специфический для ядрышек белок фибрилларин (В-36, мол. масса 34 кДа) располагается в ПФК, где он осуществляет процессинг пре-рРНК в комплексе с другими РНП, в состав которых входит U3 мяРНК, необходимая для начального этапа процессинга 45S рРНК. Фибрилларин обнаруживается также в остаточных ядрышках — в «ядрышковом матриксе».
Белок С23 (110 кДа), или «нуклеолин», локализуется в зоне плотного фибриллярного компонента и в фибриллярных центрах ядрышек, но также и в зонах ядрышковых организаторов митотических хромосом. Следовательно, он обнаруживается как на транскрибируемых, так и на неактивных участках рибосомных генов. В препаратах распластанных ядрышек он выявляется над транскрипционными единицами («елочками»). Он обнаружен во фракциях, содержащих предшественники рибосом. Функции его до конца не ясны, хотя стало известно, что белок С23 может играть важную структурную роль в процессе транскрипции: своим N-концом, на котором находятся лизиновые группы, он связывается с ядрышковым хроматином, а С-концом — с транскрибируемым спейсером (tsi) на 45S рРНК. Обнаружено, что этот белок связывается не с ДНК транскрипционной единицы, а с ДНК, имеющей нуклеосомное строение (вероятно, со спейсерными участками).
Белок В-23 (нуклеофозин, мол. масса 37 кДа) с помощью иммуноцитохимических методов локализован в области ПФК и главным образом в зоне гранулярного компонента. Считается, что В-23 участвует в промежуточных и терминальных стадиях биогенеза рибосом и в транспорте пре-рибосом.
Общая схема работы ядрышка как специального локуса синтеза рибосом
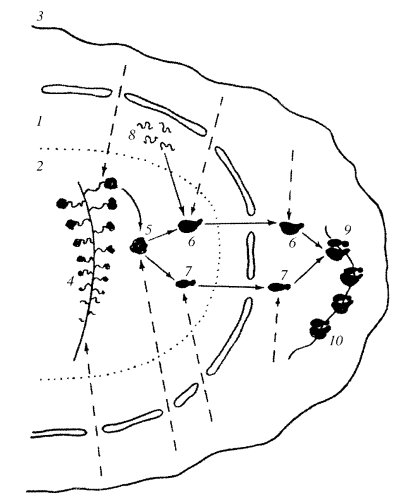
При становлении синтеза рРНК в ядрышках на поверхности ФЦ происходит активация транскрипционных единиц — связывание с факторами транскрипции и РНК-полимеразой I, которая начинает считывать первичный транскрипт рРНК. По мере прохождения первой РНК-полимеразы I на освобождающийся участок транскрипционной единицы садится следующая РНК-полимераза и начинается синтез новой рРНК. Одновременно и последовательно на одном р-гене могут находиться до сотни РНК-полимераз I, от которых отходят транскрипты разной степени завершенности. Конечным продуктом является пре-рРНК или 45S рРНК. По мере синтеза растущие цепи рРНК одеваются рибосомными белками, поступающими в ядро из цитоплазмы, так что сразу образуются цепи РНП-предшественников. Совокупность продуктов транскрипции нескольких транскрипционных единиц образует вокруг ФЦ зону ПФК. Конечным продуктом такого синтеза является рибонуклеопротеидный тяж, или глобула, имеющая константу седиментации около 80S и содержащая одну молекулу 45S рРНК. После отделения 45S рРНК в терминальной точке транскрипционной единицы происходит расщепление — процессинг 45S рРНК, в конце которого образуются 40S и 60S рибосомные субъединицы. Синтез малых субъединиц в ядрышке занимает примерно 30 мин, а больших - около 1 ч. В ядрышке незрелая 60S рибосомная субъединица, кроме двух фрагментов рРНК (28S и 5,8S), связывается с третьим (5S), который синтезировался независимо от хромосом с ядрышковыми организаторами на других хромосомах. Такие новообразованные рибосомные субъединицы особым образом выходят из ядра в цитоплазму через ядерные поры. В цитоплазме такие незрелые рибосомы могут связаться с дополнительными белками. 40S субъединица сначала связывается с иРНК и только затем с большой 60S субъединицей, образуя полную 80S функционирующую рибосому (рис. 92).
|
Рис. 92. Общая схема работы ядрышка 1 — ядро; 2 — область ядрышка; 3 — плазматическая мембрана клетки; 4 — транскрипционная единица; 5 — 80S РНП, содержащая 45S пре-рРНК; 6 — большая рибосомная субъединица; 7 — малая рибосомная субъединица; 8 — 5S рРНК, синтезируемая вне ядрышка; 9 — полная 80S работающая рибосома; 10 — полирибосома. Пунктирные стрелки - потоки белков из цитоплазмы к рРНК |
Новые, неканонические функции ядрышек
Последние данные показывают, что кроме синтеза рРНК ядрышко участвует во многих других аспектах экспрессии генов.
Первые намеки (1965 г.) на признаки полифункциональности ядрышек были получены при изучении гетерокарионов. Так, при слиянии человеческих клеток HeLa с эритроцитами кур были получены гетерокарионы с первоначально совершенно разными ядрами. Ядра клеток HeLa были функционально активны, в них шел синтез разнообразных РНК. Исходные ядра эритроцитов кур содержали сверхконденсированный хроматин, не содержали ядрышек и не транскрибировались. В гетерокарионе после слияния с клетками HeLa в ядрах эритроцитов кур хроматин начинал деконденсироваться, активировалась транскрипция, появлялись ядрышки. Иммуноцитохимическими методами изучалось появление в гетерокарионах белков, характерных для куриных клеток. Несмотря на то что в клетках HeLa имелась готовая система функционирования рибосом и были сформированы ядрышки, появление куриных белков откладывалось до тех пор, пока не возникали ядрышки в ядрах эритроцитов. Это означало, что ядрышко куриного эритроцита как-то должно вовлекаться в образование куриных иРНК, т.е. ядрышко должно играть какую-то роль в продукции куриных иРНК.
Позднее были накоплены данные в поддержку этой гипотезы. Было обнаружено, что созревание (сплайсирование) с-mус иРНК в клетках млекопитающих происходит в ядрышках. В ядрышках были выявлены сплайсосомные малые РНК (sn РНК), факторы сплайсинга пре-иРНК.
Далее в ядрышках обнаруживаются РНК, входящие в SRP-частицы, участвующие в синтезе белков в эндоплазматическом ретикулуме. С ядрышком оказалась ассоциирована РНК теломеразы - рибонуклеопротеида (обратная транскриптаза). Имеется много данных о локализации в ядрышках процессинга малых ядерных РНК, входящих в состав сплайсосом, и даже о процессинге тРНК.
Ядрышко во время митоза: периферический хромосомный материал
С помощью световой микроскопии ядрышко выявляется во время интерфазы, в митотических клетках оно исчезает. При использовании цейтраферной микрокиносъемки можно наблюдать в живых клетках как по мере конденсации хромосом в интерфазе происходит исчезновение ядрышка. Сначала оно слегка уплотняется, но затем ко времени разрыва ядерной оболочки начинает быстро терять плотность, становится рыхлым и на глазах быстро исчезает, как бы тает. При этом видно, что часть ядрышкового материала растекается между хромосомами. В метафазе и анафазе ядрышки как таковые отсутствуют. Первые признаки новых ядрышек появляются после средней телофазы, когда уже достаточно разрыхлились хромосомы дочерних ядер, имеющие новую ядерную оболочку. В это время вблизи деконденсирующихся хромосом появляются плотные тельца — предъядрышки. Обычно их число выше, чем число ядрышек в интерфазе. Позднее, уже в G1-периоде клеточного цикла предъядрышки растут, начинают объединяться друг с другом, их общее число падает, но суммарный объем возрастает. Общий объем ядрышка удваивается в S-G2-фазах. В некоторых случаях в профазе (культуры клеток человека) при конденсации хромосом крупные ядрышки распадаются на более мелкие, которые в митозе исчезают.
На самом деле никакого полного исчезновения, или «растворения», ядрышка нет: происходит изменение его структуры, редукция одной части его компонентов при сохранении другой. Так, было показано, что аргентофильные гранулы в интерфазных ядрышках, обнаруживаемые в световом микроскопе, начинают в профазе сливаться друг с другом, одновременно уменьшаясь в объеме, минимальный размер они занимают в метафазе, локализуясь в зонах ядрышковых организаторов хромосом. В таком виде они существуют до средней телофазы, когда выявляются в виде отдельных множественных предъядрышек, разбросанных среди деконденсированных хромосом. Уже в конце телофазы такие аргентофильные предъядрышки начинают расти. Таким образом, можно видеть, что во время митоза исчезновению подвергается только часть ядрышкового компонента, в то время как аргентофильный компонент сохраняется, постоянно существует во время митоза и переносится на хромосомах в дочерние ядра.
Радиоавтографическими исследованиями показано, что исчезновение ядрышек совпадает с прекращением синтеза клеточной (в основном рибосомной) РНК, который возобновляется в поздней телофазе, совпадая по времени с появлением новых ядрышек.
Кроме того, обнаружено, что активность РНК-полимеразы I также исчезает на средних стадиях митоза. Это дало основание считать, что новообразование ядрышек связано с восстановление синтеза рРНК в дочерних клетках.
Но в то же время существуют факты, указывающие на перманентное, постоянное присутствие ядрышковых компонентов в течение всего клеточного цикла. Это в первую очередь относится к аргентофильному материалу ядрышек.
Цитологи в начале XX века часто наблюдали во время митоза появление какого-то нехроматинового материала, окружающего каждую хромосому. Этот материал, или «матрикс», митотических хромосом, как считали, мог иметь ядрышковое происхождение, и его роль могла заключаться в том, что он может служить источником новых ядрышек в дочерних ядрах после митоза.
Электронная микроскопия показала, что «матрикс» - нехроматиновый компонент митотических хромосом, состоящий из скопления рыхло расположенных фибрилл и гранул, имеющих рибонуклеопротеидную природу, морфологически сходных с компонентами, входящими в состав интерфазных ядрышек, выявляется в условиях конденсации митотических хромосом как растительного, так и животного происхождения. При этом некоторые компоненты ядрышек диссоциируют и уходят в цитоплазму (большая часть РНП-частиц), в то время как другие тесно связываются с поверхностью хромосом, образуя основу «матрикса» или, как этот компонент теперь называют, основу периферического хромосомного материала (ПХМ) (рис. 93). Этот фибриллярногранулярный материал, синтезированный до митоза, переносится хромосомами в дочерние клетки. В ранней телофазе еще в отсутствие синтеза РНК по мере деконденсации хромосом происходит структурное перераспределение компонентов ПХМ. Его фибриллярные компоненты начинают собираться в мелкие ассоциаты - предъядрышки, которые могут сливаться друг с другом, собираться в зоне ядрышкового организатора хромосом в поздней телофазе, где возобновляется транскрипция рРНК.
|
Рис. 93. Микрофотография периферического хромосомного материала (ПХМ) в делящихся клетках эндосперма пшеницы, полученная с помощью электронного микроскопа (фото В.В. Буракова) а — метафаза, каждая хромосома окружена ПХМ; б — то же при окраске на РНП по Бернхарду. 1 — хромосомы; 2 — ПХМ; 3 — микротрубочки веретена |
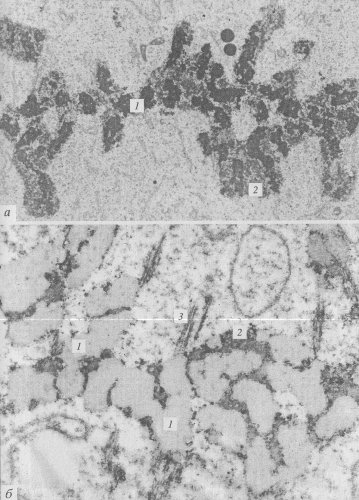
Новый этап в изучении периферического материала митотических хромосом связан с использованием иммуноцитохимических методов выявления ядрышковых белков. Было показано, что митотические хромосомы действительно участвуют в переносе в дочерние клетки белков ядрышек, белков ядерного остова и различных РНП. Например, установлено, что ядрышковые белки, участвующие в транскрипции рРНК (РНК-полимераза I, топоизомераза I, фактор инициации транскрипции UBF и др.), аккумулируются в зоне ядрышкового организатора, в то время как белки, связанные с процессингом пре-рРНК (фибрилларин, нуклеолин, В-23), а также некоторая часть пре-рРНК и малые ядрышковые РНП переносятся поверхностью хромосом в составе периферического хромосомного материала (рис. 94).
|
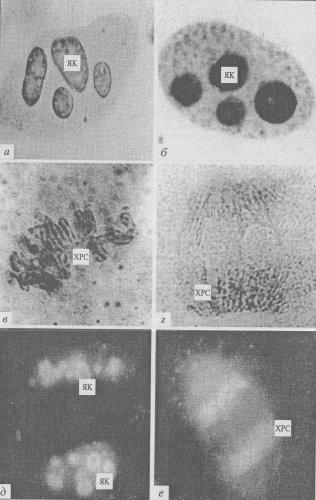
Рис. 94. Микрофотография периферического хромосомного материала в делящихся клетках эндосперма пшеницы, полученная с помощью светового микроскопа (фото Е.М. Лазаревой). а-г — окраска AgNO3 : а — интерфаза, 6 — ранняя профаза, в — метафаза, г — поздняя анафаза; д,е — окраска антителами к ядрышковому белку фибрилларину: д — ранняя интерфаза, е — анафаза. ЯК — ядрышки; ХРС — хромосомы |
Кроме того, в состав ПХМ могут входить некоторые негистоновые белки из состава ядерного интерфазного остова (рис. 95).
|
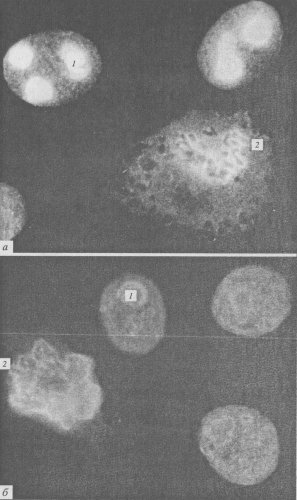
Рис. 95. Микрофотография периферического хромосомного материала (ПХМ) в клетках СПЭВ, полученная с помощью светового микроскопа (фото М.И. Мурашовой) а — окраска флуоресциируюшими антителами к ядрышковому белку В-23; б — окраска антителами к белку 40 кДа ядерного матрикса. 1 — ядрышки в интерфазных клетках; 2 — поверхностное окрашивание хромосом (ПХМ) |
Следовательно, митотические хромосомы не только выполняют свою главную функцию - переносят генетический материал в виде ДНК, но, кроме того, участвуют в переносе целого ряда белков и РНК (рис. 96).
|
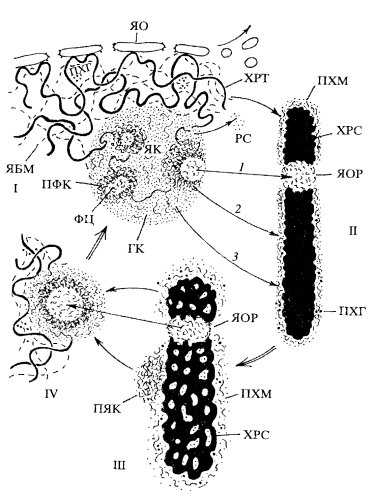
Рис. 96. Участие периферического хромосомного материала (ПХМ) в переносе хромосомами компонентов ядрышка и ядерного белкового матрикса на разных стадиях ядерного цикла I — интерфаза; II — метафаза; III — телофаза; IV — ранняя интерфаза. 1 — компоненты транскрипционного комплекса синтеза рРНК (рДНК, РНК-полимераза I, факторы транскрипции); 2 — компоненты процессипга рРНК.; 3 — предшественники рибосом и белки матрикса ядрышка. ЯК — ядрышко; ЯОР — ядрышковый организатор хромосомы; ФЦ — фибриллярный центр; ПФК — плотный фибриллярный компонент; ГК — гранулярный компонент ядрышка; PC — рибосомы; ЛЯК — предъядрышки; ХРТ — хроматин; ХРС — хромосома; ПХГ — периферические гранулы; ЯБМ — ядерный белковый матрикс; ЯО — ядерная оболочка |
Биологический смысл появления ПХМ на поверхности митотических хромосом может заключаться в том, что переносимые хромосомами белки не являются случайными «пассажирами», а представляют собой комплекс белков разного происхождения: ферменты и факторы ядрышковой транскрипции, процессинга рРНК, сборки рибосом, незрелые предшественники рибосом и, кроме того, белки ядерного и ядрышкового матрикса, также содержащие малые ядерные РНП и все компоненты, связанные с образованием нерибосомных РНК, с их сплайсингом и др. Другими словами, ПХМ переносит в новые ядра многие белковые компоненты и ферменты, что создает условия, необходимые для форсированного возобновления синтеза и созревания рибосом, а также синтеза информационных РНК. Митотическая хромосома переносит в новое ядро не только генетическую информацию в виде ДНК хроматина, но и необходимые компоненты синтетического аппарата, готового к активации транскрипции в новом клеточном цикле. Хромосома при клеточном делении «все свое несет с собой», как гласит латинская поговорка.
Величко В.В. © Copyright 2008.![]()
![]()