 О проектеПредмет клеточнойСтроение и химияЦитоплазмаСистемыЦитоскелетМеханизмы
О проектеПредмет клеточнойСтроение и химияЦитоплазмаСистемыЦитоскелетМеханизмыЦЕНТРАЛЬНАЯ ДОГМА МОЛЕКУЛЯРНОЙ БИОЛОГИИ
ХРОМОНЕМНЫЙ (ЧЕТВЕРТЫЙ) УРОВЕНЬ УПАКОВКИ ХРОМАТИНА
НЕРИБОСОМНЫЕ ПРОДУКТЫ КЛЕТОЧНОГО ЯДРА
ЯДЕРНАЯ ОБОЛОЧКА

Исследуя структурную организацию хроматина и хромосом, можно определенно говорить о нескольких уровнях компактизации ДНК.
Первый — нуклеосомный, дающий семикратное уплотнение ДНК и составе фибрилл ДНП, второй — фибрилла диаметром 30 нм, или нуклеомерный уровень, с 40-70-кратной степенью упаковки, третий — доменно-петлевой, или хромомерный, приводящий к 600-700-кратному уплотнению ДНК в составе этих структур. Для поддержания первых двух уровней компактизации было достаточно участия только гистоновых белков, тогда как петлевые и розеткоподобные доменные структуры уже требовали участия негистоновых белков и перехода от спирального, или соленоидного, типа укладки ДНК к образованию компактных глобулярных структур, состоящих из петель хроматиновых фибрилл диаметром 30 нм, к структурам типа хромомеров, имеющих уже размеры 0,1-0,2 мкм.
Однако еще в классических работах цитологов начала XX века как в интерфазных ядрах, так и, особенно, в митотических хромосомах описывались нитчатые структуры — хромонемы, имеющие толщину 0,1—0,2 мкм. Их удавалось наблюдать как на фиксированных объектах, так и в живых клетках. Подробные исследования ультраструктуры митотических хромосом на разных этапах митоза с помощью электронной микроскопии полностью подтвердили наличие этого четвертого уровня компактизации хроматина (рис. 71).
|
Рис. 71. Хромонемы на ультратонких срезах а — профазные хромосомы пиона; б — телофазные хромосомы клеток культуры ткани |
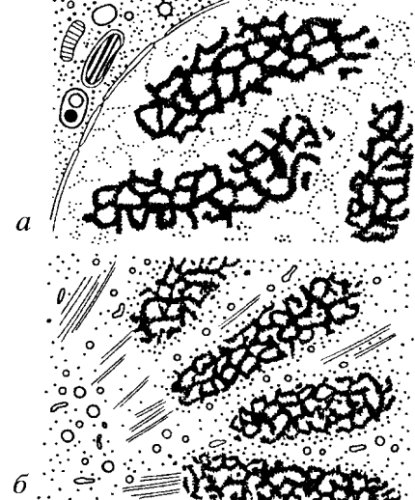
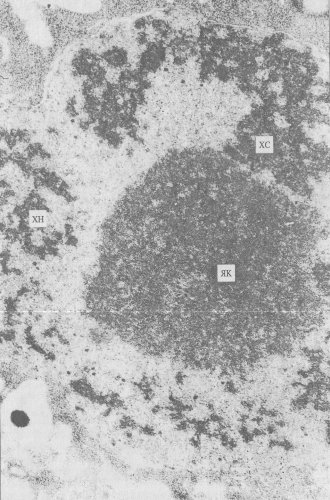
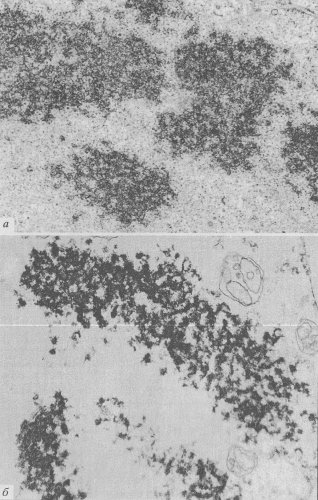
При изучении ультраструктурных основ строения митотических хромосом необходимо учитывать хромонемный уровень компактизации хроматина. Хромонему — нитчатую хроматиновую структуру со средней толщиной 0,1—0,2 мкм удается проследить в естественных условиях на разных стадиях начальной конденсации хромосом в профазе митоза и при деконденсации хромосом в телофазе. Причем такие хромонемы выявляются как в клетках растений, так и животных (рис. 72 и 73).
|
Рис. 72. Хромонемная организация хромосом Раеоnica arborea в профазе митоза ЯК — ядрышко; ХС — хромосома; ХН — хромонемы |
|
Рис. 73. Хромонемная организация хромосом а — участок хромосом клеток СПЭВ в ранней телофазе (фото В.Ю. Полякова); 6 — хромонемы в составе экспериментально деконденсированных хромосом (фото В.В. Буракова) |
Изучение профазных хромосом как животных, так и растений показывает, что процесс конденсации хромосомного материала включает в себя промежуточный этап - образование из фибрилл ДНП нитчатых хромонемных структур, являющихся единицей последующей хромосомной структуризации.
В естественных условиях в составе метафазных хромосом хромонемные элементы на ультратонких срезах не выявляются. Но по мере деконденсации митотических хромосом в поздней анафазе и ранней телофазе снова можно видеть признаки хромонемной организации хромосом. В поздней анафазе, когда хромосомы достигают противоположных полюсов клетки, в их структуре снова выявляются хроматиновые нитчатые образования с толщиной 0,2 мкм. При этом вся структура хромосом разрыхляется, что отражает начало общей деконденсации митотических хромосом. Эта начальная стадия деконденсации связана не с разрыхлением фибрилл ДНП внутри хромонем, а с расхождением, обособлением участков хромонемы друг от друга. Особенно заметным и выраженным этот процесс становится в телофазе. В это время хромосомы начинают увеличиваться в объеме, при этом расстояние между отдельными участками хромонемы также возрастает. В расположении отдельных нитей хромонемы, так же как и в профазных хромосомах, улавливаются признаки спиральности в их укладке: часто видны кольчатые или петлистые незамкнутые участки, иногда располагающиеся параллельно друг другу. Спиральность хромонемы в составе митотических хромосом удается наблюдать в ряде случаев при частичной искусственной деконденсации выделенных митотических хромосом (рис. 74). В поздней телофазе хромосомы уже полностью окружены ядерной оболочкой. Хромонемные элементы расходятся на значительные расстояния, но все же зоны отдельных хромосом еще выявляются. В это время некоторые участки хромонем начинают разрыхляться, их толщина возрастает. Таким образом, наблюдая за состоянием структуры и расположением хромонемных участков в ядрах и хромосомах в телофазе, можно видеть картину, обратную той, что наблюдалась в профазе: разрыхление хромосом за счет первоначального расхождения участков хромонемы и последующего их разрыхления, деконденсации самих хромонем.
|
Рис. 74. Спиральность выделенных хромосом после обработки 0,6М КСl |

Ультраструктурная организация хромонемного уровня упаковки ДНП хорошо выявляется при постепенном экспериментальном разрыхлении хромосом при понижении концентрации двухвалентных катионов. Оказалось, что плотное тело митотических хромосом сначала разрыхляется, благодаря чему выявляется его хромонемная организация: на срезах видно, что хромосомы представлены сечениями толстых (0,1—0,2 мкм) хромосомных нитей — хромонем (см. рис. 76). Затем, при последующем снижении концентрации двухвалентных катионов, происходит как бы распад хромонемных элементов на множество линейно расположенных глобулярных блоков хроматина с диаметром около 0,1-0,2 мкм. В дальнейшем эти блоки (хромомеры) начинают деконденсироваться: на их периферии видны петли фибрилл ДНП, а в центре остается тело хромомера. Возникает розеткоподобная структура. Важно отметить, что расположение зон с розеткоподобными хромомерами совпадает с рисунком G-бэндирования хромосом. По мере дальнейшей деконденсации петли увеличиваются в длину, а центральные участки хромомеров прогрессивно уменьшаются. При полной деконденсации все тело хромосомы представлено на срезах равномерно расположенными фибриллами ДНП.
Необходимо отметить, что в современных молекулярно-биологических исследованиях строения хромосом хромонемный уровень, как один из высших уровней упаковки ДНП, совершенно выпадает из поля зрения исследователей. Лишь в последнее время некоторые исследователи на основании косвенных данных приходят к выводам о наличии в интерфазных ядрах хромонемоподобных структур.
Общая организация митотических хромосом
Интенсивное изучение ультраструктуры хромосом началось в середине 1950-х годов, что было связано с внедрением в цитологию метода электронной микроскопии. Однако вклад электронной микроскопии в изучение структуры интерфазных и митотических хромосом оказался неизмеримо ниже того, что дал этот метод для изучения структуры цитоплазмы. Наши представления о структурной организации даже элементарных компонентов ядра и о структуре хромосом очень скудны и противоречивы. Разрыв между успехами в биохимическом изучении процессов биосинтеза ДНК и РНК, с одной стороны, и чрезвычайно медленным прогрессом в исследовании тонкой организации клеточного ядра - с другой, объясняется многими причинами. Одна из основных причин та, что современные методы не позволяют изучать ядро и хромосомы в целостной совокупности составляющих их элементов. Хромосома оказалась слишком мала для детального анализа с помощью светового микроскопа и слишком велика и плотна для изучения в электронном микроскопе. На выделенных хромосомах в электронном микроскопе не удается выявить все детали из-за наложений проекций разных уровней и можно наблюдать лишь характер формы или же тонкую структуру в ограниченных участках. Исследование ультратонких срезов хромосом ограничивается характеристикой отдельных элементов без возможности получить объемное представление о всей структуре. Это происходит из-за того, что в данном случае мы можем исследовать лишь плоские сечения, составляющие только 0,05-0,025-ную часть общего объема ядра. Так как для ядра и хромосом характерно преобладание тонких и длинных спутанных фибриллярных структур, которые на ультратонких срезах будут иметь вид беспорядочно разбросанных коротких отрезков, то по таким сечениям воссоздать трехмерную картину взаимосвязи этих элементов друг с другом практически невозможно.
При изучении ультраструктуры хромосом исследователи сталкиваются с парадоксальной ситуацией: чем ближе подходим к высшим структурным уровням организации хромосом, тем меньшей по объему и более низкой по надежности становится информация об этой важнейшей клеточной структуре (рис. 75).
|
Рис. 75. Общий вид хромосом а — хромосомы клеток культуры СПЭВ на ультратонком срезе; 6 — вид выделенных метафазных хромосом в просвечивающем электронном микроскопе; в — вид выделенных метафазных хромосом в сканирующем электронном микроскопе |

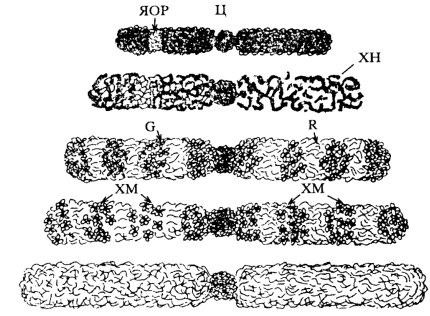
Действительно, получена полная информация о генетическом коде человека, хорошо изучен нуклеосомный уровень компактизации ДНК, определен общий петлевой доменный характер дальнейшей укладки ДНК, подтверждаются представления о хромонемном уровне, но все же на сегодня мы до конца не знаем, как построена митотическая хромосома (рис. 76).
|
Рис. 76. Ступени дифференциальной деконденсации хромосом Ц — центромера; ЯОР — ядрышковый организатор; G, R — сегменты хромосомы; ХМ — хромомеры; ХН — хромонема |
Трудности изучения хромосом связаны кроме всего прочего с тем, что это очень лабильная структура, легко меняющая свою морфологию в зависимости от условий эксперимента. Так, обращает на себя внимание свойство митотических хромосом обратимо изменять свой объем при изменении ионного окружения. Как уже указывалось, применение гипотонических растворов приводит к набуханию хромосом, но при возвращении их в изотонические условия они вновь приобретают исходную морфологию. Из этого следует, что существует какойто механизм, стабилизирующий общую организацию хромосомы. В хромосоме существует какой-то структурный порядок, алгоритм взаимодействия компонентов, который инвариантно приводит интерфазную развернутую, деконденсированную хромосому в состояние плотного тела (митотическая хромосома), не меняющего ни своей толщины, ни длины, ни особенностей структуры в бесчисленном ряду клеточных поколений.
Как уже говорилось, для изучения ультраструктуры хромосом широко применяется метод получения целых выделенных митотических хромосом. На таких препаратах видно, что в состав хромосом входят элементарные фибриллы диаметром 25—30 нм. Однако уловить характер их укладки, какой-либо порядок в их расположении не удается. Хромосомы в этом случае имеют вид тел, состоящих как бы из перепутанных изгибающихся фибрилл, или, по образному выражению одного из цитологов, напоминают результат аварии на макаронной фабрике.
На препаратах таких выделенных и распластанных хромосом нет реальной возможности выяснить, из какого числа нитей состоит хромосома, тем более проследить путь и порядок укладки одной нити от начала до конца, если бы она была основой хромосомы. Более того, процесс выделения хромосом приводит к изменению их структуры. Легко видеть в световом микроскопе, что при переносе живых делящихся клеток в гипотонические растворы их хромосомы резко набухают, становятся плохо различимы, увеличиваются в объеме, их оптическая плотность уменьшается. В целом митотические хромосомы в этих условиях ведут себя так же, как препараты выделенного хроматина: набухают и переходят в менее конденсированное состояние. Такое воздействие на них гипотонической среды приводит к потере субструктуризации хромосомы.
Однако о том, что такая субструктуризация существует, говорит масса фактов не только электронно-микроскопических, но и полученных с помощью светового микроскопа. Вся совокупность морфологических и биохимических данных должна быть учтена при воссоздании трехмерной организации митотических хромосом.
В 1970-х годах удалось уловить общий принцип структурной организации митотической хромосомы. Было обнаружено, что хромосомы не теряют своей морфологической целостности, не распадаются даже при резком набухании, вызванном удалением всех гистонов. Это достигается обработкой выделенных хромосом растворами полианионов, декстрансульфата и гепарина. В этом случае хромосомы настолько деконденсируются, что перестают быть видны в фазово-контрастном микроскопе. При добавлении же флуорохрома, связывающегося с ДНК (этидиум бромид), было видно, что сильно набухшие хромосомы не разваливались, а значительно (до 4 раз) увеличивались в длину и ширину. Такие сильно набухшие, лишенные гистонов хромосомы помещали на подложку и рассматривали с помощью электронного микроскопа.
Оказалось, что набухшие хромосомы состоят из двух компонентов: из рыхлой сети плотных фибрилл в центральных участках (хромосомный остов - скэффолд), повторяющих контуры метафазных хромосом (осевые компоненты), и из многочисленных длинных тонких петель, отходящих от них в поперечном направлении (рис. 77). Была показана белковая природа осевых компонентов и ДНК в составе петель. Средний размер боковых петель составлял около 30 мкм. Если такие препараты обработать ДНКазой, то можно получить белковые остовы и анализировать их состав. Оказалось, что в них присутствует около 20 белков негистоновой природы, сходных с белками ядерного матрикса. Исходя из этого, была предложена модель структурной организации митотических хромосом. В ее основе лежит принцип поперечного расположения петель ДНК вдоль белковой осевой структуры. В принципе этот тип организации митотической хромосомы очень напоминает хромосомы типа «ламповых щеток», встречающихся в процессе мейоза.
|
Рис. 77. Латеральные петли ДНК и осевые компоненты метафазной хромосомы после полной экстракции гистонов |
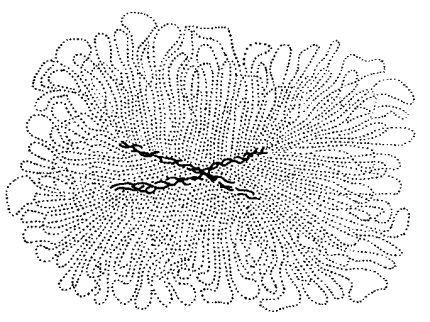
Петлевое расположение ДНК вдоль хромосомы получило в дальнейшем целый ряд подтверждений. Однако при разных способах депротеинизации кроме петель в периферии набухших хромосом можно было выявить и розеткоподобные структуры, состоящие из ДНК.
В последнее время получены данные, говорящие о том, что осевые структуры могут представлять собой артефакт, получившийся в результате монтажа и высушивания дегистонизированных хромосом на подложке. На самом же деле в теле хромосомы существуют негистоновые белковые связки (скрепки), сшивающие основания боковых петель ДНК, но эти связки разбросаны рыхло по объему хромосомы (рис. 78). Как бы то ни было, принцип петлевой поперечной укладки ДНК в теле хромосомы очень важен для понимания ее общей ультраструктурной организации.
|
Рис. 78. Схема возможного расположения петель ДНК (1) и белковых «скрепок» (2), связь между которыми определяет параметры хромосом |
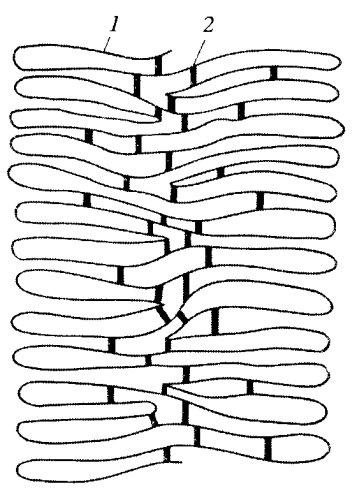
Необходимо подчеркнуть, что на поперечных и продольных сечениях митотических хромосом, фиксированных в нативном состоянии, внутри клеток никаких центральных или осевых элементов не обнаружено. Они выявляются только после удаления из выделенных хромосом всего набора гистонов, чему предшествует изоляция хромосом в гипотонической среде.
На основании этих наблюдений широкое распространение нашла христоматийная схема, объясняющая общую структуру митотической хромосомы (рис. 79). По этой схеме первым уровнем компактизации ДНК является нуклеосомная фибрилла толщиной 10 нм, где вокруг одной нуклеосомы оборачивается 146 п.н. ДНК с коэффициентом компактности (к.к.), равным 6—7; второй уровень — фибрилла (соленоид) толщиной 30 нм имеет к.к., равный 40; третий уровень — петлевой домен (60 т.п.н.) размером 0,2—0,3 мкм имеет к.к., равный 680. Далее отрезок примерно с 18—20 петлевыми доменами образует вокруг осевого элемента хромосомы один виток диаметром 0,7-0,8 мкм (толщина хроматиды) с коэффициентом компактизации 12·104. Такой виток из петлевых доменов может представлять собой минимального размера бэнд, а набор из нескольких витков — средний бэнд.
|
Рис. 79. Схема уровней организации хроматид (Альбертс и др., 1994) |
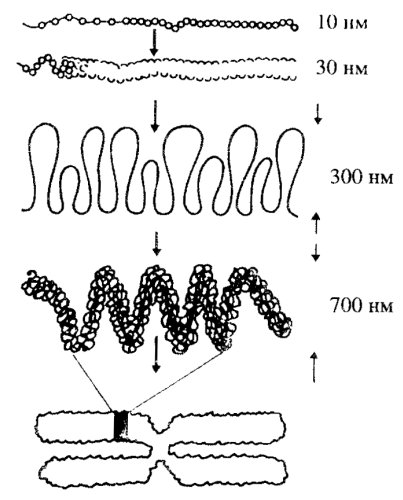
По другим представлениям можно предположить, что петле ДНК, выявляемой на хромосомах, лишенных гистонов, соответствует хромомер, промежуточным этапом деконденсации которого является розеткоподобная структура ДНП.
Важно отметить, что хромомерные участки ДНП встречаются и в интерфазных ядрах (там они называются хромоцентрами). Оказалось, что порядок их деконденсации такой же, что и в митотической хромосоме: из хромоцентров возникают розеткоподобные структуры с петлями ДНП по их периферии.
Итак, можно несколько иначе оценить некоторые этапы компактизации ДНК, которые приводят в конце концов к построению плотного тела митотической хромосомы (рис. 80).
|
Рис. 80. Схема различных уровней компактизации хроматина 1 — нуклеосома; 2 — нуклеомер, «сверхбусина»; 3 — хромомер, петлевой домен; 4 — хромонема; 5 — хроматида. Стрелка показывает, что петля домена (3) представлена фибриллой толщиной 30 нм (2) |
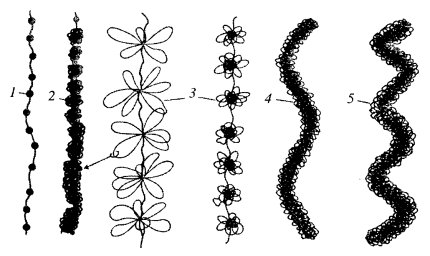
Первый уровень — нуклеосомный — образует сверхскручивание ДНК по поверхности гистоновой сердцевины. Второй — нуклеомерный (сверхбусина), где идет объединение 8-10 нуклеосом в виде глобулы. Так как все эти уровни компактизации происходят на огромных линейных молекулах ДНК, то ряд сближенных нуклеомеров образует фибриллу ДНП диаметром 30 нм. Третий уровень — хромомерный: петли фибрилл ДНП, объединенные скрепками из негистоновых белков, образуют компактные тела, которые при искусственной деконденсации дадут розетковидные структуры. Расположение петлевых доменов — хромомеров, может быть неравномерным; участки тела митотической хромосомы, обогащенные ими, могут соответствовать бэндам или сегментам при дифференциальной окраске хромосом. Четвертый уровень — хромонемный: сближенные в линейном порядке хромомеры образуют толстые (0,1-0,2 мкм) хромосомные нитчатые структуры, которые уже можно наблюдать и в световом микроскопе. Характер упаковки этой нити в теле хроматиды еще недостаточно выяснен; возможна спиральная укладка хромонемы, но не исключено образование ею и еще одного уровня петлистых структур.
Конечно, такая общая схема организации митотических хромосом очень неполно отражает особенности строения их специализированных участков, таких как ядрышковый организатор, теломеры и центромеры.
Величко В.В. © Copyright 2008.![]()
![]()