 О проектеПредмет клеточнойСтроение и химияЦитоплазмаСистемыЦитоскелетМеханизмы
О проектеПредмет клеточнойСтроение и химияЦитоплазмаСистемыЦитоскелетМеханизмыМИКРОТРУБОЧКИ

Общие свойства
Одним из обязательных компонентов цитоскелета эукариот являются микротрубочки (рис. 265). Это нитчатые неветвящиеся структуры толщиной 25 нм, состоящие из белков-тубулинов и ассоциированных с ними белков. Тубулины микротрубочек при полимеризации образуют полые трубки, откуда и их название. Длина их может достигать нескольких микрометров; самые длинные микротрубочки встречаются в составе аксонемы хвостов спермиев.
|
Рис. 265. Микротрубочки фибробласта, окрашенные антителами к тубулину (фото А.В. Буракова) Я — ядро; КЦ — клеточный центр |
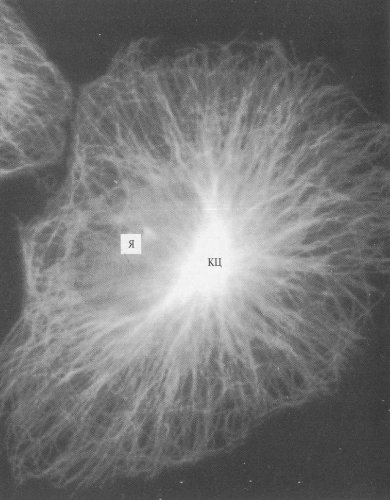
Микротрубочки обнаруживаются в цитоплазме интерфазных клеток, где они располагаются поодиночке или небольшими рыхлыми пучками, или в виде плотноупакованных микротрубочек в составе центриолей, базальных телец и в ресничках и жгутиках. При делении клеток большая часть микротрубочек клетки входит в состав веретена деления.
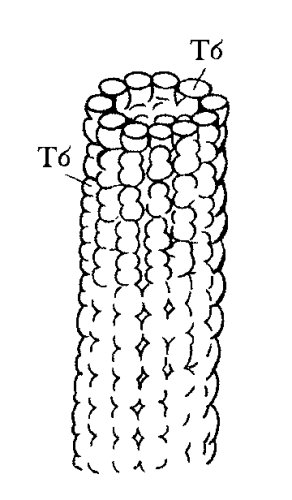
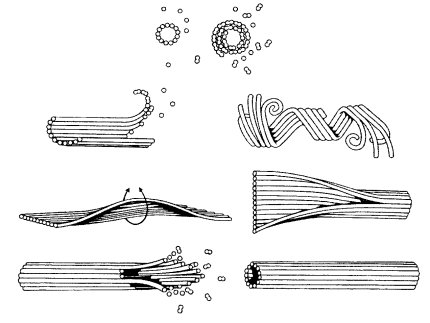
В морфологическом отношении микротрубочки представляют собой длинные полые цилиндры с внешним диаметром 25 нм (рис. 266). Стенка микротрубочек состоит из полимеризованных молекул белка тубулина. При полимеризации молекулы тубулина образуют 13 продольных протофиламентов, которые скручиваются в полую трубку (рис. 267). Размер мономера тубулина составляет около 5 нм, равного толщине стенки микротрубочки, в поперечном сечении которой видны 13 глобулярных молекул.
|
Рис. 266. Строение микротрубочки Тб — субъединица, димер тубулина |
|
Рис. 267. Стадии самосборки микротрубочек |
Молекула тубулина представляет собой гетеродимер, состоящий из двух разных субъедниц: из α-тубулина и β-тубулина, которые при ассоциации образуют собственно белок тубулин, изначально поляризованный. Обе субъединицы мономера тубулина связаны с ГТФ, однако на α-субъдинице ГТФ не подвергается гидролизу, в отличие от ГТФ на β-субъединице, где при полимеризации происходит гидролиз ГТФ до ГДФ. При полимеризации молекулы тубулина объединяются таким образом, что с β-субъединицей одного белка ассоциируется α-субъединица следующего белка и т.д. Следовательно, отдельные протофибриллы возникают как полярные нити, и соответственно вся микротрубочка тоже является полярной структурой, имеющей быстро растущий плюс-конец и медленно растущий минус-конец (рис. 268).
|
Рис. 268. Полимеризация тубулинов На плюс-конце микротрубочки происходит включение димеров тубулинов, связанных с ГТФ, на минус-конце преобладает диссоциация тубулинов |
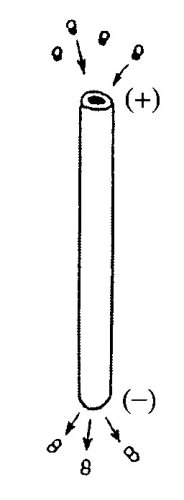
При достаточной концентрации белка полимеризация происходит спонтанно. При спонтанной полимеризации тубулинов осуществляется гидролиз одной молекулы ГТФ, связанной с β-тубулином. Во время наращивания длины микротрубочки связывание тубулинов идет с большей скоростью на растущем плюс-конце. Но при недостаточной концентрации тубулина микротрубочки могут разбираться с обоих концов. Разборке микротрубочек способствуют понижение температуры и наличие ионов Са2+.
Существует ряд веществ, которые влияют на полимеризацию тубулина. Так, алкалоид колхицин, содержащийся в безвременнике осеннем (Colchicum autumnale), связывается с отдельными молекулами тубулина и предотвращает их полимеризацию. Это приводит к падению концентрации свободного тубулина, способного к полимеризации, что вызывает быструю разборку цитоплазматических микротрубочек и микротрубочек веретена деления. Таким же действием обладают колцемид и нокодозол, при отмывании которых происходит полное восстановление микротрубочек.
|
Сеть микротрубочек разрушена обработкой нокодазолом (A). Затем клетки инкубировались в среде, свободной от нокодозола и фиксировались через 2 (B), 4 (C), 6 (D), 8 (E) и 10 (F) минут. Клетки окрашены антителами к β-тубулину. Некоторые микротрубочки полимеризовались на центросоме, но большинство образовалось в цитоплазме. Масштаб — 10 мкм. |
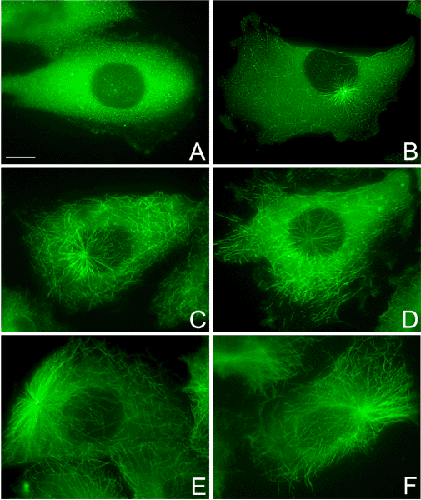
Стабилизирующим действием на микротрубочки обладает таксол, который способствует полимеризации тубулина даже при его низких концентрациях.
Все это показывает, что микротрубочки являются очень динамичными структурами, которые могут достаточно быстро возникать и разбираться.
В составе выделенных микротрубочек обнаруживаются ассоциированные с ними дополнительные белки, так называемые МАР-белки (MAP - microtubule associated proteins). Эти белки, стабилизируя микротрубочки, ускоряют процесс полимеризации тубулина (рис. 269).
|
Рис. 269. Стабилизация микротрубочек с помощью МАП |
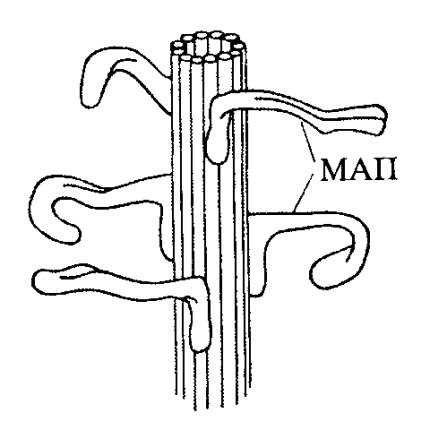
В последнее время процесс сборки и разборки микротрубочек стали наблюдать в живых клетках. После введения в клетку меченных флуорохромами антител к тубулину и при использовании электронных систем усиления сигнала в световом микроскопе можно видеть, что в живой клетке микротрубочки растут, укорачиваются, исчезают, т.е. постоянно находятся в динамической нестабильности. Оказалось, что среднее время полужизни цитоплазматических микротрубочек со
ставляет всего лишь 5 мин. Так, за 15 мин около 80% всей популяции микротрубочек обновляется. При этом отдельные микротрубочки могут на растущем конце медленно (4—7 мкм/мин) удлиняться, а затем достаточно быстро (14-17 мкм/мин) укорачиваться. В живых клетках микротрубочки в составе веретена деления имеют время жизни около 15—20 с. Считается, что динамическая нестабильность цитоплазматических микротрубочек связана с задержкой гидролиза ГТФ, это приводит к тому, что на плюс-конце микротрубочки образуется зона, содержащая негидролизованные нуклеотиды («ГТФ-колпачок»). В этой зоне молекулы тубулина связываются с большим сродством друг к
другу, и, следовательно, скорость роста микротрубочки возрастает. Наоборот, при потере этого участка, микротрубочки начинают укорачиваться.
Однако 10-20% микротрубочек остаются относительно стабильными достаточно долгое время (до нескольких часов). Такая стабилизация наблюдается в большой степени в дифференцированных клетках. Стабилизация микротрубочек связана или с модификацией тубулинов или с их связыванием с дополнительными (MAP) белками микротрубочек и с другими клеточными компонентами.
Ацетилирование лизина в составе тубулинов значительно увеличивает стабильность микротрубочек. Другим примером модификации тубулинов может быть удаление терминального тирозина, что также характерно для стабильных микротрубочек. Эти модификации обратимы.
Сами микротрубочки не способны к сокращению, однако они являются обязательными компонентами многих движущихся клеточных структур, таких как реснички и жгутики, как веретено клетки во время митоза, как микротрубочки цитоплазмы, которые обязательны для целого ряда внутриклеточных транспортов, таких как экзоцитоз, движение митохондрий и др.
В целом же роль цитоплазматических микротрубочек может быть сведена к двум функциям: скелетной и двигательной. Скелетная, каркасная, роль заключается в том, что расположение микротрубочек в цитоплазме стабилизирует форму клетки; при растворении микротрубочек клетки, имевшие сложную форму, стремятся приобрести форму шара. Двигательная роль микротрубочек заключается не только в том, что они создают упорядоченную, векторную, систему движения. Микротрубочки цитоплазмы в ассоциации со специфическими ассоциированными моторными белками образуют АТФазные комплексы, способные приводить в движение клеточные компоненты.
Практически во всех эукариотических клетках в гиалоплазме можно видеть длинные неветвящиеся микротрубочки. В больших количествах они обнаруживаются в цитоплазматических отростках нервных клеток, в отростках меланоцитов, амеб и других изменяющих свою форму клетках (рис. 270). Они могут быть выделены сами или же можно выделить их образующие белки: это те же тубулины со всеми их свойствами.
|
Рис. 270. Расположение микротрубочек в цитоплазме фибробласта (а), меланоцита (б) и нейрона (в) |
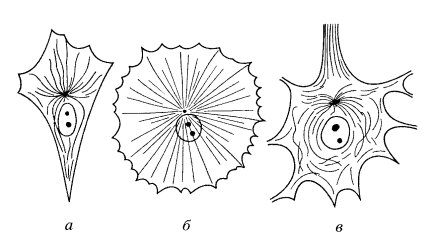
Центры организации микротрубочек
Рост микротрубочек цитоплазмы происходит полярно: наращивается плюс-конец микротрубочки. Так как время жизни микротрубочек очень коротко, то должно постоянно происходить образование новых микротрубочек. Процесс начала полимеризации тубулинов — нуклеация, происходит в четко ограниченных участках клетки, в так называемых центрах организации микротрубочек (ЦОМТ). В зонах ЦОМТ осуществляется закладка коротких микротрубочек, обращенных своими минус-концами к ЦОМТ. Считается, что в зонах ЦОМТ минус-концы заблокированы специальными белками, предотвращающими или ограничивающими деполимеризацию тубулинов. Поэтому при достаточном количестве свободного тубулина будет наращиваться длина микротрубочек, отходящих от ЦОМТ. В качестве ЦОМТ в клетках животных участвуют главным образом клеточные центры, содержащие центриоли, о чем будет сказано позже. Кроме того, в качестве ЦОМТ может служить ядерная зона и во время митоза полюса веретена деления.
Наличие центров организации микротрубочек доказывается прямыми экспериментами. Так, если в живых клетках полностью деполимеризовать микротрубочки или с помощью колцемида или путем охлаждения клеток, то после снятия воздействия первые признаки появления микротрубочек будут проявляться в виде радиально расходящихся лучей, отходящих от одного места (цитастер). Обычно у клеток животного происхождения цитастер возникает в зоне клеточного центра. После такой первичной нуклеации микротрубочки начинают отрастать от ЦОМТ и заполнять всю цитоплазму. Следовательно, растущие периферические концы микротрубочек будут всегда плюс-концами, а минус-концы будут располагаться в зоне ЦОМТ (рис. 271 и 272).
|
Рис. 271. Динамика восстановления микротрубочек (МТ) а — исходный фибробласт; б — исчезновение МТ после охлаждения клетки; в — появление первых микротрубочек в виде цитастера в зоне клеточного центра; г — восстановление цитоплазматических микротрубочек |
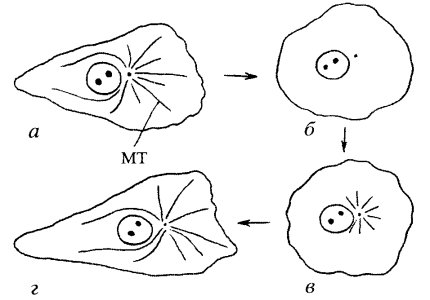
|
Рис. 272. Полярность в расположении микротрубочек (МТ) в цитоплазме Все плюс-концы МТ направлены от центра к периферии клетки |
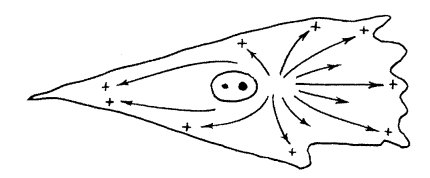
Цитоплазматические микротрубочки возникают и расходятся от одного клеточного центра, с которым многие теряют связь, могут быстро разбираться или, наоборот, стабилизироваться при ассоциации с дополнительными белками.
Одно из функциональных назначений микротрубочек цитоплазмы заключается в создании эластичного, но одновременно устойчивого внутриклеточного скелета, необходимого для поддержания формы клетки. У дисковидных по форме эритроцитов амфибий по периферии клетки лежит жгут циркулярно уложенных микротрубочек; пучки микротрубочек характерны для различных выростов цитоплазмы (аксоподии простейших, аксоны нервных клеток и т.д.).
Действие колхицина, вызывающего деполимеризацию тубулинов, сильно меняет форму клетки. Так, если отросчатую и плоскую клетку в культуре фибробластов обработать колхицином, то она теряет полярность. Точно таким же образом ведут себя другие клетки: колхицин прекращает рост клеток хрусталика, отростков нервных клеток, образование мышечных трубок и т.д. Так как при этом не исчезают элементарные формы присущего клеткам движения, такие как пиноцитоз, ундулирующие движения мембран, образование мелких псевдоподий, то роль микротрубочек заключается в образовании каркаса для поддержания клеточного тела, для стабилизации и укрепления клеточных выростов. Кроме того, микротрубочки участвуют в процессах роста клеток. Например, у растений в процессе растяжения клеток, когда за счет увеличения центральной вакуоли происходит значительный рост объема клеток, большие количества микротрубочек появляются в периферических слоях цитоплазмы. В этом случае микротрубочки, так же как и растущая в это время клеточная стенка, как бы армируют, механически укрепляют цитоплазму.
Создавая такой внутриклеточный скелет, микротрубочки могут быть факторами ориентированного движения внутриклеточных компонентов, задавать своим расположением пространства для направленных потоков разных веществ и для перемещения крупных структур. Так, в случае меланофоров (клетки, содержащие пигмент меланин) рыб при росте клеточных отростков гранулы пигмента передвигаются вдоль пучков микротрубочек. Разрушение микротрубочек колхицином приводит к нарушению транспорта веществ в аксонах нервных клеток, к прекращению экзоцитоза и блокаде секреции. При разрушении микротрубочек цитоплазмы происходит фрагментация и разбегание по цитоплазме аппарата Гольджи, разрушение митохондриального ретикулума.
Динеины и кинезины - моторные белки
Долгое время считалось, что участие микротрубочек в движении цитоплазматических компонентов заключается лишь в том, что они создают систему упорядоченного движения. Иногда в популярной литературе цитоплазматические микротрубочки сравнивают с железнодорожными рельсами, без которых движение поездов невозможно, но которые сами по себе ничего не двигают. Одно время предполагали, что двигателем, локомотивом, может быть система актиновых филаментов, но оказалось, что механизм внутриклеточного перемещения различных мембранных и немембранных компонентов связан с группой иных белков.
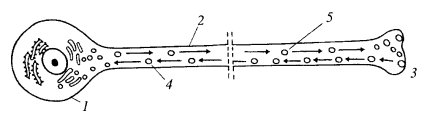
Прогресс был достигнут при изучении так называемого аксонального транспорта в гигантских нейронах кальмара. Аксоны — отростки нервных клеток, могут иметь большую длину и заполнены большим числом микротрубочек и нейрофиламентов. В аксонах живых нервных клеток можно наблюдать перемещение различных мелких вакуолей и гранул, которые двигаются как от тела клетки к нервному окончанию (антероградный транспорт), так и в противоположном направлении (ретроградный транспорт). Если аксон перетянуть тонкой лигатурой, то такой транспорт приведет к скоплению мелких вакуолей по обе стороны от перетяжки. Вакуоли, двигающиеся антероградно, содержат различные медиаторы, в том же направлении могут двигаться и митохондрии. Ретроградно двигаются вакуоли, образовавшиеся в результате эндоцитоза при рециклировании мембранных участков. Эти движения происходят с относительно высокой скоростью: от тела нейрона — 400 мм в сутки, в направлении к нейрону — 200—300 мм в сутки (рис. 273).
|
Рис. 273. Схема движения частиц в аксоне 1 — тело нейрона; 2 — аксон; 3 — нервное окончание; 4 — ретроградный транспорт; 5 — антероградный транспорт |
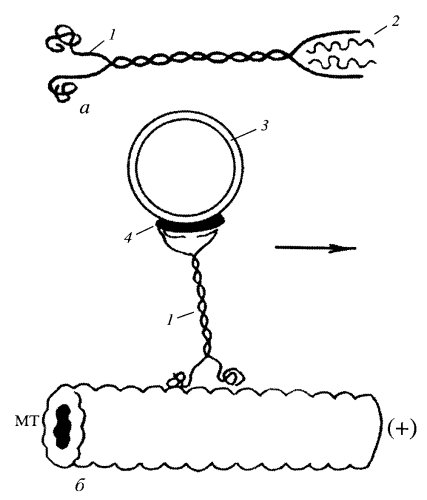
Оказалось, что из отрезка гигантского аксона кальмара можно выделить аксоплазму, содержимое аксона. В капле выделенной аксоплазмы продолжается движение мелких вакуолей и гранул. С помощью видеоконтрастного устройства можно видеть, что движение мелких пузырьков происходит вдоль тонких нитчатых структур, вдоль микротрубочек. Из этих препаратов были выделены белки, ответственные за движение вакуолей. Один из них кинезин, белок с молекулярной массой около 300 тыс. Он состоит из двух сходных тяжелых полипептидных цепей и нескольких легких. Каждая тяжелая цепь образует глобулярную головку, которая при ассоциации с микротрубочкой обладает АТФазной активностью, в то время как легкие цепи связываются с мембраной пузырьков или других частиц (рис. 274). При гидролизе АТФ изменяется конформация молекулы кинезина и генерируется перемещение частицы в направлении к плюс-концу микротрубочки. Оказалось возможным приклеить, иммобилизовать молекулы кинезина на поверхности стекла; если к такому препарату в присутствии АТФ добавить свободные микротрубочки, то последние начинают двигаться. Наоборот, можно иммобилизовать микротрубочки, но если добавить к ним мембранные пузырьки, связанные с кинезином, пузырьки начинают двигаться вдоль микротрубочек.
|
Рис. 274. Внутриклеточное перемещение вакуоли с помощью кинезина в направлении к плюс-концу микротрубочки (МТ) а — молекула кинезина: 1 — тяжелые цепи, 2 — легкие цепи; б — участие кинезина в транспорте: 1 — кинезин, 3 — мембранный груз, 4 — кинектин |
Существует целое семейство кинезинов, обладающих сходными моторными головками, но отличающихся хвостовыми доменами. Так, цитозольные кинезины участвуют в транспорте по микротрубочкам везикул, лизосом и других мембраных органелл. Многие из кинезинов связываются специфически со своими грузами. Некоторые участвуют в переносе только митохондрий, другие — только синаптических пузырьков. Кинезины связываются с мембранами через мембранные белковые комплексы — кинектины. Кинезины веретена деления участвуют в образовании этой структуры и в расхождении хромосом.
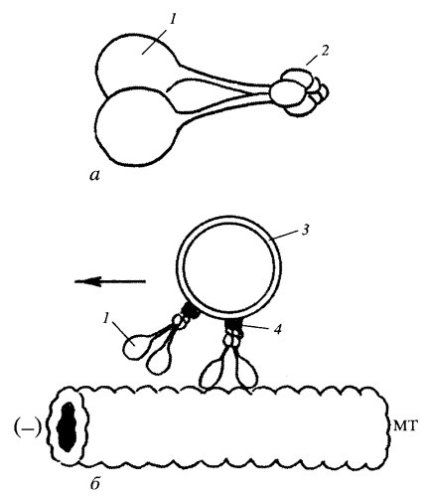
За ретроградный транспорт в аксоне отвечает другой белок — цитоплазматический динеин (рис. 275). Он состоит из двух тяжелых цепей — головок, взаимодействующих с микротрубочками, нескольких промежуточных и легких цепей, которые связываются с мембранными вакуолями. Цитоплазматический динеин является моторным белком, переносящим грузы к минус-концу микротрубочек. Динеины также делятся на два класса: цитозольные, участвующие в переносе вакуолей и хромосом, и аксонемные, отвечающие за движение ресничек и жгутиков.
|
Рис. 275. Внутриклеточное перемещение вакуоли с помощью цитоплазматического динеина к минус-концу микротрубочки (МТ) а — молекула динеина: 1 — тяжелые цепи, 2 — легкие цепи; б — участие динеина в транспорте: 1 — динеин, 3 — мембранный груз, 4 — динактин |
Цитоплазматические динеины и кинезины были обнаружены практически во всех типах клеток животных и растений.
Таким образом, и в цитоплазме движение осуществляется по принципу скользящих нитей, только вдоль микротрубочек перемещаются не нити, а короткие молекулы — «движители», связанные с перемещающимися клеточными компонентами. Сходство с актомиозиновым комплексом этой системы внутриклеточного транспорта заключается в том, что образуется двойной комплекс (микротрубочка + «движитель»), обладающий высокой АТФазной активностью.
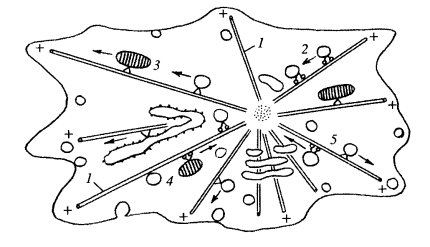
Как мы видим, микротрубочки образуют в клетке радиально расходящиеся поляризованные фибриллы, плюс-концы которых направлены от центра клетки к периферии. Наличие же плюс- и минус-направленных моторных белков (кинезинов и динеинов) создает возможность для переноса в клетке её компонентов как от периферии к центру (эндоцитозные вакуоли, рециклизация вакуолей ЭПР и аппарата Гольджи и др.), так и от центра к периферии (вакуоли ЭПР, лизосомы, секреторные вакуоли и др.) (рис. 276). Такая полярность транспорта создается за счет организации системы микротрубочек, возникающих в центрах их организации, в клеточном центре.
|
Рис. 276. Ориентация движения внутриклеточных мембранных компонентов 1 — микротрубочки; 2, 4 — движение к центру клетки, к минус-концам МТ за счет динеинов; 3, 5 — движение к периферии клетки, к плюс-концам МТ за счет кинезинов |
Величко В.В. © Copyright 2008.![]()
![]()