 О проектеПредмет клеточнойСтроение и химияЦитоплазмаСистемыЦитоскелетМеханизмы
О проектеПредмет клеточнойСтроение и химияЦитоплазмаСистемыЦитоскелетМеханизмыПРОМЕЖУТОЧНЫЕ ФИЛАМЕНТЫ

Промежуточные филаменты (ПФ) строятся из фибриллярных мономеров. Поэтому основная конструкция промежуточных филаментов напоминает канат, имеющий толщину около 8—10 нм. Они локализуются главным образом в околоядерной зоне и в пучках фибрилл, отходящих к периферии клеток и располагающихся под плазматической мембраной (рис. 238, 240 и 241). Встречаются промежуточные филаменты во всех типах клеток животных, но особенно они обильны в тех клетках, которые подвержены механическим воздействиям: клетки эпидермиса, нервные отростки, гладкие и исчерченные мышечные клетки. В клетках растений ПФ не обнаружены.
|
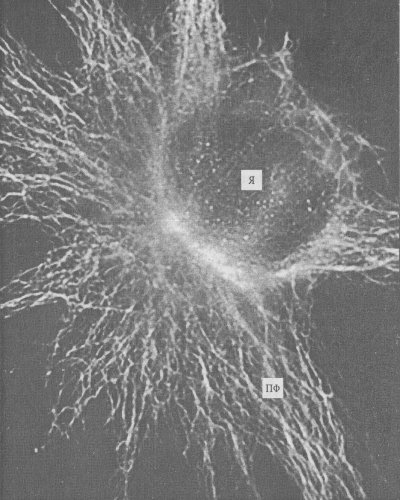
Рис. 240. Промежуточные филаменты (ПФ) фибробласта в культуре ткани, окрашенные флуоресцирующими антителами к виментину (фото Л.А. Горгидзе) Я — ядро |
|
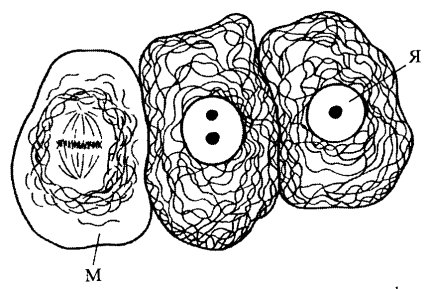
Рис. 241 . Расположение промежуточных филаментов в клетках эпителия М — митоз; Я — ядро |
В состав промежуточных филаментов входит большая группа изобелков (родственных белков), которую можно разделить на четыре типа. Первый тип составляют кератины, кислые и нейтральные, встречающиеся в эпителиальных клетках; они образуют гетерополимеры из этих двух подтипов. Кератины, кроме того, имеют, некоторую гетерогенность, зависящую от тканевого источника. Так, в эпителиях встречается до 20 форм кератинов, 10 форм других кератинов найдено в волосах и ногтях. Молекулярная масса кератинов колеблется от 40 до 70 тыс.
Второй тип белков ПФ включает в себя три вида белков, имеющих сходную молекулярную массу (45—53 тыс.). Это — виментин, характерный для клеток мезенхимного происхождения, входящий в состав цитоскелета клеток соединительной ткани, эндотелия, клеток крови. Десмин характерен для мышечных клеток, как гладких, так и исчерченных. Глиальный фибриллярный белок входит в состав ПФ некоторых клеток нервной глии — в астроциты и некоторые шванновские клетки. Периферинвходит в состав периферических и центральных нейронов.
Третий тип — белки нейрофиламентов (молекулярная масса от 60 до 130 тыс.), встречается в аксонах нервных клеток.
И наконец, четвертый тип — белки ядерной ламины. Хотя эти последние имеют ядерную локализацию, они сходны по строению и свойствам со всеми белками промежуточных филаментов.
Как уже говорилось, промежуточные филаменты построены из фибриллярных белков наподобие каната. При этом некоторые белки могут образовывать сополимеры, например виментин с десмином или виментин с глиальными белками.
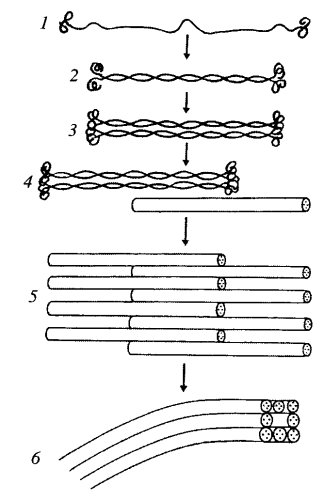
Все белки промежуточных филаментов обладают сходной аминокислотной последовательностью из 130 остатков в центральной части фибриллярной молекулы, которая обладает α-спиральным строением. Концевые же участки молекул имеют разные последовательности аминокислот, разную длину и не имеют α-спирального строения. Наличие протяженных α-спиральных участков позволяет двум молекулам образовывать двойную спираль, подобно тому как это происходит в молекуле миозина, что приводит к образованию палочковидного димера длиной около 48 нм. Два димера, объединяясь бок о бок, образуют короткий протофиламент — тетрамер, толщиной около 3 нм. Такие протофиламенты могут объединяться в более толстые и длинные фибриллы, и в конечном итоге образуется промежуточный полный филамент, состоящий из восьми продольных протофиламентов (рис. 242).
|
Рис. 242. Этапы полимеризации белков (1—5) и промежуточных филаментов 1 — отдельная молекула; 2 — димер; 3 — тетрамер-протофиламент; 4, 5 — полимеризация протофиламентов; 6 — сформированный промежуточный филамент |
Иначе полимеризуются белки ядерной ламины: они образуют димеры с головками на одном конце и полимеризуются, создавая рыхлую прямоугольную решетку. Такие слои ламины быстро разрушаются во время митоза при фосфорилировании ламинов.
Цитоплазматические промежуточные филаменты относятся к самым стабильным и долгоживушим элементам цитоскелета. Однако in vivo наблюдается включение инъецированных меченых молекул кератина в состав ПФ эпителиальных клеток. ПФ устойчивы к действию солей низкой и высокой концентрации, разрушаются только после воздействия денатурирующих растворов, таких как мочевина.
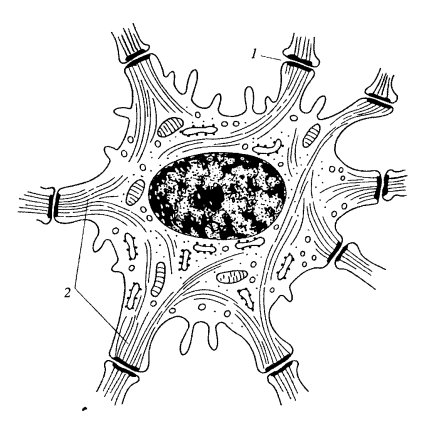
Такая структура и химическая устойчивость промежуточных филаментов, вероятно, определяют и их физическую устойчивость. Они служат как бы истинно опорной системой в клетках, подвергающихся значительным физическим нагрузкам. В клетках кожного эпидермиса промежуточные филаменты образуют пучки (тонофиламенты), связанные с десмосомами, и создают жесткую внутриклеточную сеть (рис. 243). Так, в нервных аксонах, простирающихся на многие десятки сантиметров, ПФ, или нейрофиламенты, создают жесткую основу, обеспечивающую гибкость и целостность тонких цитоплазматических отростков нервных клеток. В поперечно исчерченных мышечных клетках десминовые филаменты входят в состав Z-дисков и связывают их друг с другом как в составе саркомера, так и в соседних миофибриллах, а также с плазматической мембраной.
|
Рис. 243. Клетка шиповатого слоя эпидермиса кожи 1 — десмосомы; 2 — кератиновые промежуточные филаменты |
Специфических ингибиторов полимеризации белков промежуточных филаментов пока еще не найдено. Поэтому остается неясным сам процесс сборки и разборки этих элементов цитоскелета в живой клетке. Вероятнее всего, что они подобно ламинам деполимеризуются при действии цитоплазматических киназ, приводящих к их фосфорилированию. Выделенные промежуточные филаменты под действием фосфорилаз могут распадаться на мономеры, деполимеризоваться.
Топографически в клетке расположение промежуточных филаментов повторяет расположение микротрубочек, они как бы идут бок о бок. При разрушении микротрубочек колхицином происходит так называемый коллапс промежуточных филаментов: они собираются в плотные пучки или кольца вокруг ядра. Восстановление новой сети промежуточных филаментов начинается от зоны клеточного центра. Это наводит на мысль, что центром их полимеризации или нуклеации могут быть центры, общие с микротрубочками.
Величко В.В. © Copyright 2008.![]()
![]()