 О проектеПредмет клеточнойСтроение и химияЦитоплазмаСистемыЦитоскелетМеханизмы
О проектеПредмет клеточнойСтроение и химияЦитоплазмаСистемыЦитоскелетМеханизмы
В предыдущих разделах уже много и часто говорилось о движении: движутся хромосомы к полюсам клетки во время митоза, перемещаются вакуоли клеточных органелл, движется клеточная поверхность. Кроме того, в клетках растений и животных наблюдаются токи цитоплазмы (например, в растительных клетках или у амебы). Более того, отдельные клетки (свободноживущие одноклеточные организмы или специфические типы клеток в многоклеточных животных организмах) обладают способностью активно перемещаться, «ползать» (рис. 237). Некоторые клетки имеют специализированные структуры, реснички или жгутики, которые позволяют им или самым перемещаться, или перемещать окружающую их жидкость. Наконец, у многоклеточных животных организмов есть специализированные клетки, мышечная работа которых позволяет производить различные движения органов, отдельных его частей и всего организма. В основе всех этих многочисленных двигательных реакций лежат общие молекулярные механизмы. Кроме того, наличие каких-либо двигательных аппаратов должно сочетаться и структурно связываться с существованием опорных, каркасных или скелетных внутриклеточных образований. Поэтому можно говорить (описывать и изучать) об опорно-двигательной системе клеток.
|
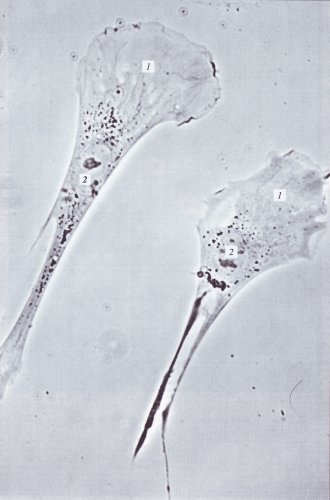
Рис. 237. Поляризованные движущиеся фибробласты в культуре ткани (фото И.С. Григорьева) 1 — ламеллоплазма; 2 — ядро |
Само понятие о цитоскелете или скелетных компонентах цитоплазмы разных клеток было высказано Н.К. Кольцовым, выдающимся русским цитологом еще в начале XX века. К сожалению, они были забыты и только в конце 1950-х годов с помощью электронного микроскопа эта скелетная система было переоткрыта.
Огромный вклад в изучение цитоскелета внес метод иммунофлуоресценции, который помог разобраться в химии и динамике этого чрезвычайно важного компонента клетки. Цитоскелетные компоненты представлены нитевидными, неветвящимися белковыми комплексами, или филаментами (тонкими нитями).
Существуют три системы филаментов, различающихся по химическому составу, ультраструктуре и функциональным свойствам. Самые тонкие нити - это микрофиламенты; их диаметр составляет около 6 нм и состоят они в основном из белка актина. К другой группе нитчатых структур относятся микротрубочки, которые имеют диаметр 25 нм и состоят в основном из белка тубулина. Третья группа представлена промежуточными филаментами с диметром около 10 нм (промежуточный по сравнению с 6 и 25 нм), образующимися из разных, но родственных белков (рис. 238 и 239).
|
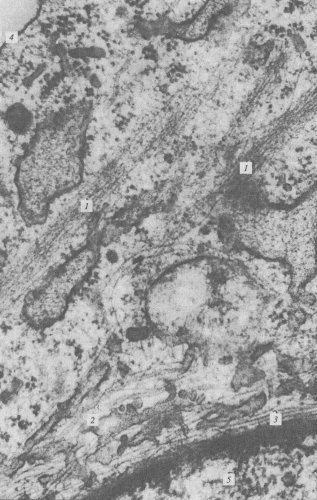
Рис. 238. Микрофотография элементов цитоскелета, полученная с помощью электронного микроскопа 1 — пучки микрофиламентов; 2 — микротрубочки; 3 — промежуточные филаменты; 4 — плазматическая мембрана; 5 — ядро |
|
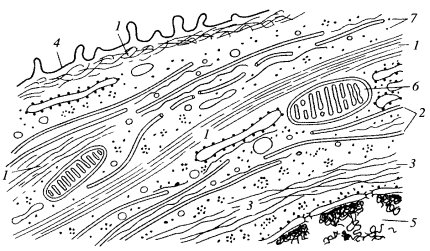
Рис. 239. Схематическое изображение цитоскелетных компонентов клеток 1 — микрофиламенты; 2 — микротрубочки; 3 — промежуточные филаменты; 4 — плазматическая мембрана; 5 — ядро; 6 — митохондрии; 7 — рибосомы |
Все эти фибриллярные структуры могут участвовать в качестве составных частей в процессе физического перемещения клеточных компонентов или даже целых клеток, кроме того, в ряде случаев они выполняют сугубо каркасную скелетную роль. Элементы цитоскелета встречаются во всех без исключения эукариотических клетках; аналоги этих фибриллярных структур встречаются и у прокариот. Степень выраженности их в разных клетках может быть различной. Например, клетки эпидермиса кожи особенно богаты промежуточными филаментами, мышечные клетки - актиновыми микрофиламентами, особенно много микротрубочек в пигментных клетках, меланоцитах, в отростках нервных клеток и т.д.
Общим для элементов цитоскелета является то, что все они представляют собой белковые, неветвящиеся фибриллярные полимеры, нестабильные, способные к полимеризации и деполимеризации. Такая нестабильность может приводить к некоторым вариантам клеточной подвижности, например к изменению формы клетки. Некоторые компоненты цитоскелета при участии специальных дополнительных белков могут стабилизироваться или образовывать сложные фибриллярные ансамбли и играть только каркасную роль. При взаимодействии с другими специальными белками-транслокаторами (или моторными белками) они могут участвовать в разнообразных клеточных движениях.
По своим свойствам и функциям элементы цитоскелета можно разделить на две группы: только каркасные фибриллы — промежуточные филаменты, а также опорно-двигательные, например актиновые микрофиламенты, взаимодействующие с моторными белками — миозинами, и тубулиновые микротрубочки, взаимодействующие с моторными белками динеинами и кинезинами.
Причем во второй группе фибрилл цитоскелета (микрофиламенты и микротрубочки) могут происходить два принципиально различных способа движения. Первый из них основан на способности основного белка микрофиламентов — актина, и основного белка микротрубочек — тубулина, к полимеризации и деполимеризации, что при связи этих белков с плазматической мембраной может вызывать ее морфологические изменения в виде образования выростов (псевдоподий и ламеллоподий) на краю клетки. Псевдоподии и тонкие выросты (филоподии) могут или втягиваться обратно в клетку, или закрепляться на поверхности клетки и затем участвовать в перемещении клетки по субстрату.
При другом способе передвижения фибриллы актина (микрофиламенты) или тубулина (микротрубочки) являются направляющими структурами, по которым перемещаются специальные подвижные белки — моторы. Последние могут связываться с мембранными или фибриллярными компонентами клетки и тем самым участвовать в их перемещении.
Величко В.В. © Copyright 2008.![]()
![]()