 О проектеПредмет клеточнойСтроение и химияЦитоплазмаСистемыЦитоскелетМеханизмы
О проектеПредмет клеточнойСтроение и химияЦитоплазмаСистемыЦитоскелетМеханизмыОБЩИЕ СВОЙСТВА БИОЛОГИЧЕСКИХ МЕМБРАН - ЛИПОПРОТЕИДНЫХ КОМПЛЕКСОВ
ВАКУОЛЯРНАЯ СИСТЕМА ВНУТРИКЛЕТОЧНОГО ТРАНСПОРТА
ГЛАДКИЙ РЕТИКУЛУМ И ДРУГИЕ МЕМБРАННЫЕ ВАКУОЛИ

Гладкий (агранулярный) эндоплазматический ретикулум
Гладкий ЭПР представляет собой часть мембранной вакуолярной системы. В морфологическом отношении он также представлен мембранами, образующими мелкие вакуоли и трубки, канальцы, которые могут ветвиться, сливаться друг с другом. В отличие от гранулярного на мембранах гладкого ЭПР нет рибосом (рис. 190). Диаметр вакуолей и канальцев гладкого ЭПР обычно около 50-100 нм. Выраженность сети из этих мембранных элементов может быть неодинаковой как для различных клеток, так и внутри одной клетки. Большей частью такие гладкие канальцы образуют скопления, или зоны. Так, в клетках эпителия кишечника гладкий ЭПР локализуется главным образом в апикальной, верхней части клетки, вблизи всасывающей поверхности. В клетках печени зоны гладкого ЭПР часто связаны с местами отложения гликогена. Встречаются клетки, где гладкий ЭПР занимает большую часть объема цитоплазмы (например, в интерстициальных клетках семенника, в растительных железистых терпеноидогенных клетках).
|
Рис. 190. Зона гладкого ретикулума в интерстициальных клетках надпочечника |

Неоднократно была установлена непрерывность перехода между гладкой формой ЭПР и гранулярной его формой. Часто можно наблюдать, как цистерна гранулярного ЭП Р теряет на своей поверхности рибосомы и становится «гладкой» (рис. 191). При этом такой участок цистерны делается неровным, начинает как бы ветвиться, переходя в трубочки и канальцы гладкого ЭПР. Этот участок часто называют переходным из-за того, что именно здесь образуются и отделяются транспортные пузырьки, переносящие новосинтезированные белки и липиды к зоне аппарата Гольджи. Гладкий ЭПР является вторичным по отношению к гранулярному ЭПР, т.е. происходит из последнего. Так, у крысенка перед рождением в печеночных клетках образуется большое количество гранулярного ЭПР, но сразу после рождения появляется масса трубочек гладкого ЭПР. Ряд биохимических, морфологических и радиоавтографических данных приводит к заключению, что гранулярный ЭПР увеличивается в объеме, растет за счет синтезирующихся мембран, которые остаются в его составе, или, потеряв рибосомы, превращается в гладкий ЭПР. Например, при использовании радиоактивных предшественников мембранных компонентов и при получении отдельных фракций гладкого и гранулярного ЭПР было обнаружено, что при интенсивном разрастании гладкого ЭПР метка вначале появляется в гранулярном ЭПР и только спустя некоторое время — в гладком ЭПР.
|
Рис. 191. Переход гранулярного эндоплазматического ретикулума в гладкий в печеночной клетке |

Несмотря на топографическую связь и общность происхождения, эти два представителя ЭПР резко отличаются друг от друга в функциональном отношении. Как уже указывалось, отсутствие рибосом на гладком ЭПР прямо говорит о его непричастности к синтезу белков. Деятельность гладкого ЭПР скорее можно связать с метаболизмом липидов и некоторых внутриклеточных полисахаридов.
Участие гладкого ЭПР в синтезе триглицеридов и липидов было показано при изучении процессов всасывания жиров клетками кишечного эпителия. В просвете кишечника жиры распадаются до жирных кислот и моноглицеридов. В апикальных участках клеток кишечника видно при этом накопление осмиофильных гранул внутри просветов канальцев гладкого ЭПР. Это связано с ресинтезом новых триглицеридов из поступивших в клетку предшественников с образованием липидов и липопротеидов, которые с помощью вакуолей аппарата Гольджи выводятся из клеток и попадают в лимфатическое русло.
Мелкие капли липидов иногда в комплексе с белками можно наблюдать и в клетках печени, причем эти капли встречаются в полостях гладкого ЭПР около зоны аппарата Гольджи. Если крысам давать вещества, приводящие к образованию отложений больших капель жира (жировая дистрофия), то первые мелкие липидные капельки появляются в гладком ЭПР, но иногда и в полостях гранулярного ЭПР.
Гладкий ЭПР особенно в большом объеме встречается в клетках, секретирующих стероиды, в частности в клетках коркового вещества надпочечника. Основные ферменты синтеза стероидов были обнаружены во фракциях микросом, образовавшихся при разрушении гладкого ЭПР из этих клеток. Гладким ретикулумом богаты интерстициальные клетки семенников, участвующие в синтезе стероидных гормонов, а также клетки сальных желез в самом начале накопления жира.
Тесная топографическая связь гладкого ЭПР с отложениями гликогена (запасного внутриклеточного полисахарида животных и грибов) в гиалоплазме различных клеток говорит о его участии в метаболизме углеводов. В клетках печени, в мышечных волокнах гликоген откладывается в зонах, свободных от гранулярных цистерн ЭПР, но богатых пузырьками и канальцами гладкого ЭПР. Такие зоны гладкого ЭПР могут увеличиваться в размере как при исчезновении гликогена (например, при голодании), так и при увеличении его отложений (рис. 192 и 193).
|
Рис. 192. Отложение гликогена (Г) в зоне гладкого эндоплазматического ретикулума (ЭПР) гепатоцита М — митохондрии |

|
Рис. 193. Зона мелких вакуолей гладкого эндоплазматического ретикулума и гранулы гликогена в клетке печени мыши В — вакуоли; Г — гликоген; М — митохондрии |

В печени часто увеличение зон гладкого ЭПР связано с рядом патологических процессов в клетках. Так, при барбитуратных отравлениях, при действии различных канцерогенов или ядовитых веществ, при действии больших доз гормональных препаратов клетки печени теряют характерную для них базофилию цитоплазмы, в них падает содержание РНК и в цитоплазме появляются оксифильные зоны. В электронном микроскопе эти зоны представлены скоплениями гладкого ЭПР. Это явление связано с тем, что в этих местах происходят процессы деградации различных вредных веществ, процессы метаболической дезактивации, которые осуществляются целым рядом окислительных ферментов, из которых наиболее известен белок, называемый цитохром Р-450. Этот белок участвует в присоединении гидроксилыюй группы к различным, потенциально опасным водонерастворимым углеводородам или к липофильным ядовитым веществам (например, четыреххлористый углерод), попадающим в мембранный бислой. Здесь же другие ферменты добавляют к этим гидроксильным группам отрицательно заряженные молекулы (сульфат, глюкуроновая кислота), что делает метаболиты или вредные липофильные вещества растворимыми в воде, из-за чего они могут выводиться из организма вместе с мочой. Разросшийся гладкий ЭПР в клетках печени после удаления токсического вещества уничтожается, вероятно, с помощью лизосом - автофагосом.
В поперечно-полосатых мышцах вакуоли и каналы гладкого ЭПР (саркоплазматический ретикулум) окружают каждую миофибриллу (рис. 194). Здесь ЭПР выполняет специальную функцию депонирования ионов кальция. В присутствии АТФ он может активно поглощать и накапливать ионы кальция, что приводит к расслаблению мышечного волокна. Белки кальциевого насоса являются интегральными белками мембран саркоплазматического ретикулума.
|
Рис. 194. Гладкий эндоплазматический ретикулум (саркоплазматический) в скелетных мышцах (по: Fawselt , McNutt , 1969) а — на срезе; б — модель. Ф — миофибриллы; М — митохондрии; ПМ — плазматическая мембрана; Т — трабекулярмые мембраны; СР — саркоплазматический ретикулум; Z — Z-полоска |

Среди высших растений гладкий ретикулум встречается в клетках тканей, участвующих в синтезе и транспорте терпенов, стероидов и липидов.
Вакуоли растительных клеток
Клетки как низших, так и высших растительных организмов содержат в цитоплазме вакуоли, несущие ряд важных физиологических нагрузок (рис. 195). У молодых клеток может быть несколько мелких вакуолей, которые по мере роста и дифференцировки клетки сливаются друг с другом и образуют одну или несколько крупных вакуолей, занимающих до 90% объема всей клетки. Эти центральные вакуоли отделены от цитоплазмы одинарной мембраной, сходной по толщине с плазмалеммой. Мембрана, ограничивающая центральные вакуоли, носит название тонопласта. Центральные вакуоли образуются из мелких пузырьков, отщепившихся от аппарата Гольджи. Такие первичные вакуоли растут в объеме, сливаются друг с другом и в конце концов образуют одну или несколько крупных вакуолей, оттесняющих цитоплазму с ядром и органоидами к периферии клетки. Полость вакуоли заполнена так называемым клеточным соком, представляющим собой водный раствор, в который входят различные неорганические соли, сахара, органические кислоты и их соли и другие низкомолекулярные соединения, а также некоторые высокомолекулярные вещества (например, белки).
|
Рис. 195. Центральные вакуоли (ЦВ) в клетке меристемы корня (а) и в мезофиле листа (б) Я — ядро; В — вакуоли; ХП — хлоропласты |
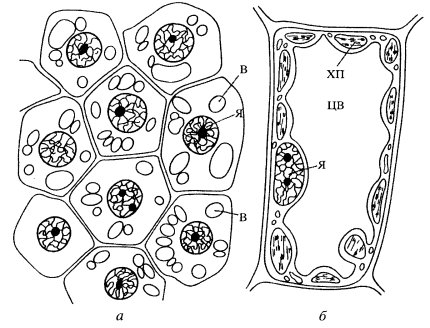
Центральные вакуоли растений выполняют многообразные и важные функции. Одной из главных их функций является поддержание тургорного давления клеток. Растворенные в соке вакуолей молекулы определяют его осмотическую концентрацию. Соответствующая молекулярная концентрация сока вакуолей и полупроницаемые свойства как ее мембраны (тонопласта), так и плазмалеммы способствуют тому, что вакуоли функционируют в качестве осмометра и придают клетке необходимую прочность и тургисцентность (напряженность).
Другая функция определяется тем, что вакуоль представляет собой большую полость, отделенную от метаболизирующей гиалоплазмы мембраной (тонопластом), обладающей свойствами полупроницаемости. Через нее, как и через плазматическую мембрану, может идти активный транспорт различных молекул. В тонопласте обнаружен АТФ-зависимый Н+-насос, направленный внутрь вакуолей и участвующий в транспорте сахаров. Поэтому вакуоли могут использоваться клетками в качестве накопительных резервуаров, где не только откладываются запасные вещества, но и собираются метаболиты, предназначенные для экскреции. Так выводятся (секретируются) из клетки все водорастворимые метаболиты. Нерастворимые в воде органические компоненты могут превращаться в растворимые глюкозиды, соединяясь с молекулами сахаров. Перечень экскретируемых в вакуоли метаболитов очень обширен. Это различные алкалоиды (например, никотин, кофеин) и полифенолы. В вакуолях происходит отложение многих глюкозидов, к которым относятся различные пигменты, например антоцианы.
Из неорганических веществ в вакуолярном соке накапливаются фосфаты калия, натрия, кальция, могут запасаться соли органических кислот (оксалаты, цитраты и др.). Это придает вакуолярному соку отчетливую кислую реакцию (рН от 2 до 5). Таким образом, можно считать, что тонопласт участвует в процессах экскреции.
Другой обширный ряд функций вакуолей связан с накоплением запасных веществ, таких как сахара и белки. Сахара в вакуолях содержатся в виде растворов, встречаются и резервные полисахариды типа инулина. В вакуолях откладываются запасные белки, что характерно для семян. Поступление белков в вакуоли, вероятнее всего, связано со способностью вакуолей ЭПР и АГ сливаться с тонопластом. Запасание белков семян злаковых происходит в так называемых алейроновых вакуолях, которые заполняются альбуминами и глобулинами, после чего вакуоли обезвоживаются, превращаясь в твердые алейроновые зерна. При прорастании семян эти зерна обводняются и снова превращаются в вакуоли. В таких новообразованных вакуолях выявляется активность некоторых ферментов: кислой фосфатазы, α-амилазы, глюкозидазы, прогеиназы и РНКазы. Следовательно, алейроновые вакуоли отчасти напоминают лизосомы, где происходит переваривание запасных белков при прорастании семян.
Гидролитические ферменты были обнаружены не только в алейроновых вакуолях, но в мелких и крупных центральных вакуолях. Наблюдалась неоднократно инвагинация (впячивание) тонопласта внутрь вакуолей, при этом часть «втянутого» материала оказывалась в полости вакуоли и там деградировала. Возможно, так выполняется аутофагическая функция вакуолей, участвующих в гидролизе дефектных клеточных компонентов. Лизосомными свойствами обладают вакуоли дрожжей. Обнаружено, что стенки вакуолей дрожжей тоже могут образовывать впячивания внутрь, затем они отщепляются от тонопласта и растворяются внутри вакуоли.
Сферосомы
Это мембранные пузырьки, встречающиеся в клетках растений. Они окрашиваются липофильными красителями, имеют высокий коэффициент преломления и поэтому хорошо видны под световым микроскопом. Сферосомы образуются из элементов эндоплазматического ретикулума. На конце цистерны ЭПР начинает накапливаться осмиофильный материал, затем от этого участка отшнуровывается и начинает расти мелкий пузырек, достигающий диаметра 0,1-0,5 мкм. Это «просферосома», окруженная одинарной мембраной. Рост сферосом и перестройка их содержимого связаны с накоплением в них масла, так что сферосома постепенно превращается в масляную каплю. Отложение липидов начинается между осмиофильными слоями мембраны. Кроме жиров в составе сферосом обнаруживают белки и среди них фермент липазу, расщепляющую липиды.
Пероксисомы (микротельца)
Это небольшие вакуоли (0,3—1,5 мкм), одетые одинарной мембраной, отграничивающей гранулярный матрикс, в центре которого располагается сердцевина, или нуклеоид (ничего не имеющий общего с нуклеоидом бактерий и вообще к ядерным структурам не относящийся).
В зоне сердцевины часто, особенно в пероксисомах печеночных клеток, видны кристаллоподобные структуры, состоящие из регулярно упакованных фибрилл, или трубочек. Изолированные сердцевины пероксисом содержат фермент уратоксидазу (рис. 196).
|
Рис. 196. Строение пероксисом в клетках печени (а) и листа табака (б) |

Пероксисомы обнаружены у простейших (амебы, тетрахимена), у низших грибов (дрожжи), у высших растений в некоторых эмбриональных тканях (эндосперм) и в зеленых частях, способных к фотореспирации. У высших позвоночных животных они найдены главным образом в печени и почках. В печени крыс на клетку число пероксисом колеблется от 70 до 100.
Пероксисомы часто локализуются вблизи мембран ЭПР. У зеленых растений пероксисомы часто находятся в тесном контакте с митохондриями и пластидами.
Впервые пероксисомы были выделены из печени и почек. Во фракциях пероксисом обнаруживаются ферменты, связанные с метаболизмом перекиси водорода. Это ферменты (оксидазы, уратоксидаза, оксидаза D-аминокислот) окислительного дезаминирования аминокислот, при работе которых образуется перекись водорода (Н2О2) и каталаза, разрушающая ее. В пероксисомах печени каталаза составляет до 40% всех белков и локализована в матриксе. Так как Н2О2 является токсическим веществом для клеток, то каталаза пероксисом может играть важную защитную роль. Пероксисомы в клетках цыплят и лягушек кроме уратоксидазы содержат ряд ферментов катаболизма пуринов.
У животных и некоторых растений (проростки клещевины) пероксисомы играют важную роль при превращении жиров в углеводы. Так, в клетках эндосперма клевещины в пероксисомах (глиоксисомах) содержатся ферменты глиоксалатного цикла.
Пероксисомы не содержат никаких нуклеиновых кислот, и все белки, из которых они состоят, кодируются ядерными генами, но их относят к саморепродуцирующимся органеллам. В пероксиомах происходит накопление специфических белков, которые синтезируются в цитозоле и имеют свои сигнальные участки. В мембране пероксисом есть рецепторный белок, который узнает транспортируемые белки. Белки мембран пероксисом, как и липиды, приходят из цитозоля. Такое накопление содержимого и рост мембраны приводят к общему росту пероксисомы, которая затем с помощью неизвестного пока механизма делится на две, т.е. самореплицируется.
Секреция белков и образование мембран у бактерий
В принципе рост плазматической мембраны и её производных у бактерий происходит тем же образом, что и образование мембран у эукариотических клеток.
Как известно, синтез белков у бактерий осуществляется на 70S рибосомах, которые так же, как и у клеток высших организмов, имеют двоякую локализацию. Большая часть рибосом бактериальных клеток образует полисомы в цитоплазме, около 25% рибосом связано с плазматической мембраной. Такие рибосомы участвуют как в синтезе белков мембраны, так и в синтезе экскретируемых белков. Многие бактериальные клетки получают питательные вещества за счет деградации полимеров около бактериальной поверхности. Для этого бактерии должны выделять гидролизирующие ферменты в окружающую среду. Это они делают намного проще, чем эукариотические клетки: часть их рибосом, локализованных на внутренней (цитоплазматической) поверхности плазматической мембраны, синтезирует белки, которые, подобно секреторным белкам, проходят через мембрану и оказываются вне клетки. Выделенные гидролазы застревают в компонентах муреиновой бактериальной стенки и там функционируют. На других рибосомах, связанных с мембранами, идет синтез белков для построения самой мембраны, подобно тому, что происходит в гранулярном ЭПР эукариотических клеток. Так что в этом отношении бактерию можно уподобить вакуоли гранулярного ЭПР, вывернутой наизнанку.
На примере бактерий хорошо изучен путь синтеза липидных компонентов мембран. Было найдено, что синтез фосфоэтидилэтаноламина происходит с помощью ферментов, являющихся интегральными белками плазматической мембраны, активные участки которых находятся на цитоплазматической стороне мембраны. Синтезированные здесь липиды встраиваются во внутренний липидный слой. Оказалось, что новосинтезированные липиды довольно быстро обнаруживаются и во внешнем слое мембраны за счет работы переносчиков - флиппаз.
Величко В.В. © Copyright 2008.![]()
![]()